La cellula
La struttura della cellula
Tutti gli esseri viventi sono costituiti da cellule, corpuscoli delimitati da un involucro e dotati di tutte le caratteristiche della vita: ogni cellula si riproduce, si nutre, respira, elimina i propri rifiuti. Per riprodursi e formare un’altra cellula identica a se stessa, la cellula contiene l’informazione genetica necessaria sotto forma di acido desossiribonucleico (DNA), organizzato in una struttura chiamata cromosoma. Per produrre le sostanze necessarie alla propria crescita e per produrre energia, la cellula funziona come un vero e proprio laboratorio chimico, presieduto da enzimi. Nonostante siano molto diverse tra loro per forma, funzione e dimensione, le cellule presentano un’organizzazione di base comune. Per questo motivo la cellula è considerata l’unità strutturale e funzionale di tutti i viventi.
La teoria cellulare

Disegno della struttura del sughero così come apparve a Robert Hooke, che la osservò in un rudimentale microscopio. Tale immagine, contenuta nella sua opera Micrographia, è all'origine dell'utilizzo del termine cellula per indicare l'unità fondamentale degli organismi viventi
La scoperta delle cellule risale al XVII secolo, all’epoca dei primi, rudimentali microscopi, ma furono necessari circa duecento anni per comprendere la loro diffusione in tutti gli esseri viventi.
Il primo scienziato che osservò una cellula fu l’inglese Robert Hooke (1635-1703), il quale nel 1665 individuò in una sezione sottile di sughero numerose cellette vuote. Hooke non collegò quelle piccole cavità all’organizzazione dei viventi, ma si limitò a dar loro un nome,
cellula. Solo nella prima metà del XIX secolo si scoprì che si compongono di cellule tutte le piante (grazie a Matthias Schleiden, nel 1838), e tutti gli animali (grazie a Theodor Schwann, nel 1839). Vent’anni più tardi, lo scienziato tedesco Rudolf Virchow (1821-1902) affermò che ogni cellula deriva da un’altra cellula preesistente. L’insieme di queste conclusioni costituisce la teoria cellulare, secondo cui:
- tutti gli organismi viventi sono costituiti da cellule;
- la cellula è l’unità strutturale e funzionale dei viventi;
- ogni cellula deriva da altre cellule.
Caratteristiche generali della cellula

Schema di una cellula animale
Viene definita cellula l’unità strutturale e funzionale degli organismi viventi o, in altri termini, il più piccolo insieme di materia dotato di “vita” (v. anche il riquadro in basso).
Ogni cellula possiede almeno tre componenti fondamentali: il materiale genetico (acido desossiribonucleico, o DNA), circondato da una matrice fluida detta citoplasma, il tutto
racchiuso da una membrana plasmatica, un sottile involucro che la separa dall’esterno.
Il numero di cellule che formano un individuo varia da 1 negli organismi unicellulari, a migliaia di miliardi negli organismi pluricellulari (1028 nell’uomo). Negli organismi unicellulari la singola cellula adempie a tutti i compiti fondamentali per la vita, in quelli pluricellulari l’insieme di cellule si differenzia, specializzandosi nello svolgimanto di parti-
colari funzioni.
Negli organismi pluricellulari le cellule hanno forma molto varia (in rapporto alla loro funzione): arrotondata, cubica, prismatica, stellata provvista di molti prolungamenti. Le dimensioni delle cellule, assai variabili, sono dell’ordine di grandezza del micron, µm (1 µm = 10-6 m) e sono in genere comprese tra 10 µm e 50 µm; le cellule più piccole sono quelle dei batteri (1 µm), mentre le più grandi sono le cellule uovo di molti animali (100 µm nell’uomo) e le fibre di alcune piante. Le ridotte dimensioni consentono alle cellule di mantenere un rapporto ottimale con l’ambiente, dal quale trarre le sostanze nutritive e nel quale scaricare le sostanze di rifiuto.
Cellule procarioti e cellule eucarioti

Differenze fondamentali tra cellula eucariote e procariote
In relazione al fatto che il materiale genetico (DNA) non formi un nucleo ben distinto oppure formi un nucleo (delimiato da una membrana), le cellule si distinguono in procarioti ed eucarioti.
Si definiscono procarioti (dal greco: nucleo primitivo) le cellule prive di una membrana che isoli il materiale genetico dal citoplasma (il DNA è semplicemente sparso nel citoplasma, concentrato in una regione chiamata nucleoide); inoltre nel citoplasma sono quasi del tutto assenti organuli e altre strutture (detti complessivamente strutture citoplasmatiche). Esternamente alla membrana plasmatica, le cellule procarioti possiedono una parete cellulare semirigida che le protegge dall’ambiente esterno. Sono organismi procarioti tutti i batteri.
Le cellule eucarioti (dal greco: nucleo ben fatto) hanno il nucleo ben distinto, racchiuso da una membrana cellulare, e numerose strutture cellulari differenziate presenti nel citoplasma. Questo tipo di cellula si trova nei protisti, nei funghi, nelle piante e negli animali.
| CELLULA PROCARIOTE | CELLULA EUCARIOTE | |
| Organismi tipici | Protisti, funghi, piante ed animali | Protisti, funghi, piante ed animali |
| Dimensioni tipiche | ~ 1-10 µm | ~ 10-100 µm (con poche eccezioni, come la cellula uovo e i neuroni motori spinali) |
| Tipo di nucleo cellulare | Nucleoide: nessun nucleo davvero definito | Nucleo racchiuso da doppia membrana |
| DNA | Cromosoma singolo, circolare, spesso presenti plasmidi in aggiunta | Cromosomi multipli, lineari, complessati da istoni |
| Sintesi di RNA e proteine | Accoppiate nel citoplasma | Sintesi dell'RNA nel nucleo e delle proteine nel reticolo endoplasmatico rugoso |
| Ribisomi | 50S+30S | 60S+40S |
| Strutture citoplasmatiche | Poche strutture | Numerose strutture racchiuse da membrane e citoscheletro |
| Movimento cellulari | Flagelli composti di flagellina | Flagelli e ciglia composte di tubulina |
| Mitocondri | Nessuno | Da uno a diverse migliaia (con alcune eccezioni) |
| Cloroplasti | Nessuno | Nelle alghe e nelle piante |
| Parete cellulare | Presente | Presente nelle piante e in alcuni funghi |
| Organizzazione | Solitamente unicellulare | Unicellulare, a colonie e in organismi pluricellulari (contenenti cellule specializzate) |
| Divisione cellulare | Scissione binaria | Mitosi (fissione o gemmazione) e meiosi |
_Tabella sulle differenza fra cellula procariote ed eucariote” >}}
Il rapporto volume-superficie nella cellula
Quando una cellula si accresce, il suo volume aumenta molto più rapidamente della superficie (per esempio, raddoppiando il raggio di una sfera, il volume aumenta di otto volte, mentre la superficie aumenta di quattro volte). Perciò, in una cellula di notevoli dimensioni, la superficie (attraverso la quale avvengono gli scambi con l’ambiente circostante) non sarebbe in grado di garantire gli scambi nutritivi adeguati alla nuova massa cellulare.
| STRUTTURA | FUNZIONE | |
| Strutture della superficie cellulare | parete cellulare* | protegge e sostiene la cellula |
| membrana plasmatica | doppio strato lipidico che isola dall'ambiente esterno | |
| giunzioni cellulari | regolano lo scambio di materiali con l'esterno | |
| plasmodesmi* | stabiliscono le comunicazioni con altre cellule | |
| Strutture in cui è organizzato il materiale genetico | cromosomi | ammassi organizzati di DNA e proteine accessorie, portatori dell'informazione generica per costruire la cellula e controllarne l'attività |
| nucleo | spazio delimitato da una membrana in cui sono contenuri i cromosomi | |
| membrana cellulare | circonda il nucleo; regola l'entrata e l'uscita di materiali dal nucleo | |
| nucleolo | sintetizza i ribosomi | |
| Strutture citoplasmatiche | citoplasma | sede dei processi metabolici, molto ricco di microtubuli e microfilamenti che mantendono la forma della celluale e partecipano attivamente ai processi di movimento |
| mitocondrio | produce energia tramite metabolismo aerobico (respirazione cellulare) | |
| cloroplasto* | realizza la fotosintesi | |
| ribosoma | sede della sintesi proteica | |
| reticolo endoplasmatico | sintetizza molte proteine (tra cui enzimi) e lipidi | |
| apparato di Golgi | modifica e smista proteine e lipidi; sintetizza carboidrati | |
| lisosoma | contiene enzimi digestivi intracellulari | |
| plastidio* | immagazzina cibo, pigmenti | |
| vacuolo | raccoglie acqua e prodotti di rifiuto: fornisce la pressione di turgore necessaria per dare sostegno alla cellila | |
| citoscheletro | dà forma e sostegno alla cellula; provvede al posizionamento e alla migrazione di parti della cellula | |
| centriolo | può produrre il fusto mitotico nelle cellule animali | |
| ciglia e flagelli | fanno muovere la cellula in un liquido o spostano il fluido sulla superficie di una cellulare ferma |
_Strutture delle cellule eucarioti animali e vegetali (*solo nelle cellule vegetali)” >}}
La membrana plasmatica
Si definisce membrana plasmatica (o cellulare) un involucro dello spessore di circa 7-8 nm (1 nm=10-9 m) che delimita la cellula, regola l’entrata e la fuoriuscita di materiali e “comunica” con le altre cellule ricevendo e producendo adeguati segnali. L’organizzazione della membrana plasmatica è descritta dal modello a mosaico fluido, elaborato nel 1972 dai biologi S.J. Singer e G.L. Nicolson. I principali costituenti della membrana plasmatica sono molecole di lipidi e di proteine; sono presenti anche i carboidrati.
Il modello a mosaico fluido della membrana plasmatica
Lipidi
I lipidi sono di tre tipi: fosfolipidi, colesterolo e glicolipidi.
I fosfolipidi si dispongono in un doppio strato a causa del loro duplice comportamento nei confronti dell’acqua: poiché la cellula è formata prevalentemente da acqua ed è immersa in un ambiente acquoso, la parte idrofila
di ogni molecola di fosfolipide (la testa) si rivolge verso il citoplasma e il liquido extracellulare, mentre la parte idrofoba (la coda) si dispone verso l’interno del doppio strato. Gli acidi grassi costituiscono le code dei fosfolipidi e rendono la membrana fluida, della stessa consistenza dell’olio; inoltre, i fosfolipidi possono spostarsi orizzontalmente nella membrana, senza però passare da uno strato all’altro. I fosfolipidi fungono da barriera sia per gli ioni sia per le molecole idrofile, che possono attraversare la membrana solo lungo specifici canali.
Il colesterolo è immerso nella membrana e conferisce al doppio strato maggiore stabilità e resistenza e al tempo stesso minore fluidità.
I glicolipidi si trovano soprattutto immersi nello strato esterno della membrana; regolano la comunicazione tra cellule.
Proteine
Numerose proteine (proteine di membrana) sono immerse nel doppio strato fosfolipidico o sono attaccate alla sua superficie (alcune proteine sono legate a carboidrati e prendono il nome di glicoproteine).
Si definiscono proteine estrinseche (o periferiche) quelle che sono inserite solo in uno dei due strati; sono dette proteine intrinseche (o integrali) le proteine completamente immerse nel doppio strato.
Come i fosfolipidi, anche le proteine sono libere di scorrere orizzontalmente nel doppio strato lipidico. Dalla diversa disposizione delle proteine sui due lati della membrana deriva l’asimmetria dei due strati. Le proteine svolgono un ruolo di catalizzatori chimici (enzimi), regolano il movimento delle molecole idrosolubili attraverso la membrana, riconoscono e legano alcune molecole del liquido extracellulare.
Carboidrati
I carboidrati sono presenti soprattutto nella membrana delle cellule eucarioti e sono legati ai lipidi (glicolipidi) o alle proteine (glicoproteine).
Il nucleo
Il nucleo è una struttura esclusiva delle cellule eucarioti. Ha forma pressoché sferica, contiene il materiale genetico (cioè l’acido desossiribonucleico, o DNA) ed è il centro di controllo, che programma e coordina le varie attività della cellula. È formato da tre componenti fondamentali: la membrana nucleare, la cromatina e uno o più nucleoli.

Schema del nucleo cellulare e del reticolo endoplasmatico annesso
La membrana nucleare separa il nucleo dal citoplasma ed è formata da due membrane a doppio strato lipidico, ciascuna perforata da sottili pori. Questi regolano il passaggio
delle grosse molecole, in particolare proteine e RNA, tra il nucleo e il citoplasma; acqua, ioni e piccole molecole possono invece attraversarli liberamente.
La materia liquida interna al nucleo è il plasma nucleare, composto da una soluzione acquosa contenente ioni, enzimi, ribosomi e prodotti intermedi della sintesi di DNA e RNA.
La cromatina è una sostanza dall’aspetto granulare, composta da lunghe molecole di DNA associate a proteine (il termine cromatina, che significa “sostanza colorata”, risale ai primi studi sul nucleo cellulare, quando, trattando la cellula con coloranti specifici, alcune regioni del nucleo risultavano maggiormente colorate). Durante la divisione cellulare (quando le informazioni genetiche sono duplicate per essere trasmesse dalla cellula madre alla cellula figlia) la cromatina si addensa e forma strutture compatte a forma di bastoncelli, i cromosomi (“corpi colorati”). Questi sono abbastanza grandi per essere osservati anche al microscopio ottico.
Nel nucleo sono presenti anche uno o più nucleoli (“piccoli nuclei”), corpuscoli di forma tondeggiante preposti alla sintesi dell’RNA ribosomiale. Questo tipo di RNA si lega a determinate proteine prodotte nel citoplasma per formare le due subunità costitutive dei ribosomi, organuli che svolgono un ruolo importante nella sintesi delle proteine. I nucleoli scompaiono durante la divisione cellulare, in cui si verifica un rallentamento della sintesi di ribosomi; quando la cellula poi ritorna alle sue funzioni abituali, il materiale del nucleolo, momentaneamente disperso nel nucleo, si organizza nuovamente in un corpuscolo sferico.
Il citoplasma
Il citoplasma è composto da una parte viscosa e molto fluida, il citosol, costituito di acqua (che rappresenta il 75-85% del peso totale della cellula), da sostanze inorganiche dissociate in forma ionica (soprattutto ioni K+ , Na+ , Ca++ e Mg++) e da diverse molecole organiche (tra cui proteine con funzione enzimatica o strutturale).
Nel citoplasma delle cellule eucarioti sono incluse numerose strutture citoplasmatiche di vario tipo, che svolgono funzioni ben precise (nei procarioti le attività cellulari sussistono, ma non sono svolte da strutture distinte). Talune di queste strutture (dette anche organuli) sono delimitate da una membrana strutturata come la membrana plasmatica, ma con modificazioni nel tipo e nel numero di fosfolipidi e proteine che permettono loro di svolgere particolari funzioni.
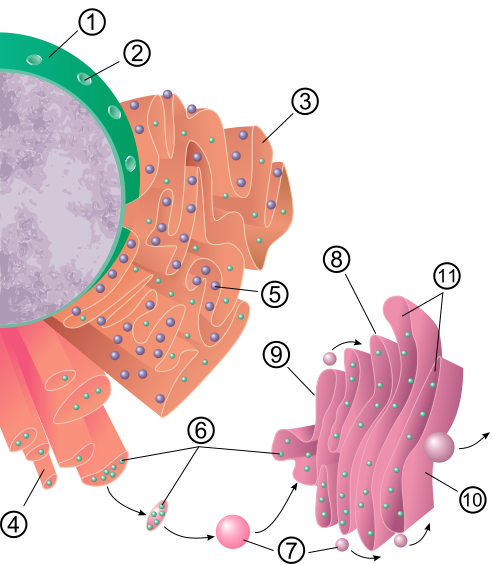
Schema del percorso secretorio tra nucleo, reticolo endoplasmatico e apparato del Golgi: 1. Membrana nucleare; 2. Poro nucleare; 3. Reticolo endoplasmatico rugoso (RER); 4. Reticolo endoplasmatico liscio (REL); 5. Ribosoma legato al RER; 6. Macromolecole; 7. Vescicole di trasporto; 8. Apparato di Golgi; 9. Faccia Cis dell'apparato di Golgi; 10. Faccia Trans dell'apparato di Golgi; 11. Cisterne del Golgi.
Nella cellula eucariote animale le strutture citoplasmatiche comprendono:
- reticolo endoplasmatico
- ribosomi
- apparato di Golgi
- lisosomi
- mitocondri
- citoscheletro
- centrioli
- ciglia e flagelli
Il reticolo endoplasmatico è una complessa rete di tubuli, sacchi e canali comunicanti, che si aprono a livello della membrana nucleare. Il reticolo endoplasmatico è di due tipi: il tipo ruvido è un proseguimento della membrana nucleare e porta sulla superficie esterna milioni di minuscoli granuli, detti ribosomi, legati alla sintesi delle proteine; il tipo liscio è privo di ribosomi ed è responsabile della sintesi dei lipidi. Lo spazio compreso tra le membrane ripiegate di entrambi i tipi di reticolo endoplasmatico è infine utilizzato per immagazzinare e trasferire molecole da un punto all’altro della cellula.
I ribosomi sono la sede della sintesi proteica, cioè l’assemblaggio degli amminoacidi per formare proteine. Sono costituiti da due subunità di dimensioni diseguali, formate da acido ribonucleico (RNA) e proteine. Possono essere associati al reticolo endoplasmatico ruvido (in questo caso sintetizzano proteine destinate a essere secrete fuori dalla cellula) oppure liberi nel citoplasma (sintetizzano proteine che la cellula trattiene al suo interno).
L‘apparato di Golgi è costituito da tubuli appiattiti al centro e rigonfi alle estremità, impilati l’uno sull’altro e terminanti a fondo cieco, dove vengono modificati e accumulati materiali vari (ormoni, proteine, lipidi), che verranno trasportati in altre parti della cellula o espulsi. A questo scopo, dalle estremità delle sacche appiattite si staccano piccole vescicole che migrano verso la membrana plasmatica e si fondono con essa; il contenuto delle vescicole viene così riversato all’esterno.
I lisosomi sono vescicole che derivano dall’apparato di Golgi e contengono enzimi idrolitici (capaci di demolire proteine e lipidi). La cellula utilizza i lisosomi per riciclare parti usurate di organuli o per “digerire” un’intera cellula (per esempio, un batterio).

Schema di un mitocondrio: 1. membrana interna, 2. membrana esterna, 3. cresta, 4 matrice
I mitocondri sono organuli tondeggianti od ovoidali, delimitati da una doppia membrana; quella interna è ripiegata su se stessa, in modo da formare dei setti (creste mitocondriali) che aumentano la superficie interna dell’organulo.
Lo spazio racchiuso dalla membrana interna è la matrice, mentre quello compreso tra le due membrane è lo spazio intermembranico. Nei mitocondri avviene la respirazione cellulare, mediante la quale viene estratta energia dalle sostanze nutritive (zuccheri e grassi) in seguito alla loro ossidazione e alla conseguente demolizione fino a diossido di carbonio e acqua. L’energia ricavata da questo processo viene immagazzinata sotto forma di molecole di ATP; al momento opportuno l’idrolisi dell’ATP rende disponibile l’energia necessaria alla cellula per svolgere le sue attività. Per questo motivo hanno un maggior numero di mitocondri le cellule con un elevato metabolismo (per esempio, le cellule muscolari); ne sono privi invece i globuli rossi. Una peculiarità dei mitocondri è la capacità di autoreplicarsi, consentita dalla presenza di un DNA mitocondriale, ribosomi e tutte le molecole necessarie alla duplicazione dell’informazione genetica. I mitocondri infatti vivono solo alcuni giorni e devono perciò essere continuamente prodotti (ciò avviene mediante il distacco dal mitocondrio primitivo di vescicole con una doppia membrana contenenti il DNA mitocondriale).
Gli organuli sono avvolti e sostenuti da fibre di natura proteica che formano una rete, il citoscheletro. Il citoscheletro inoltre fornisce sostegno alle cellule prive di una parete rigida e hanno parte attiva nella divisione cellulare e nei movimenti degli organuli e dell’intera cellula. Il citoscheletro non è una struttura rigida e permanente, poiché le fibre che lo costituiscono vengono continuamente assemblate e smontate. Queste sono distinte in tre gruppi in base alle loro dimensioni: i microfilamenti (5-7 nm di diametro), i filamenti intermedi (8-10 nm di diametro) e i microtubuli, tubuli cavi del diametro di circa 25 nm, che costituiscono anche i centrioli, le ciglia e i flagelli.
I centrioli sono strutture cave di forma cilindrica, costituiti da 9 triplette fuse di microtubuli, che si trovano in tutte le cellule animali (due per cellula) e in poche cellule vegetali. I centrioli intervengono durante la divisione cellulare per ripartire correttamente i cromosomi nelle due cellule figlie.

Ciglia dell'epitelio di un polmone (microscopio a scansione)
Le ciglia e i flagelli sono appendici filiformi e mobili con identica struttura: 9 coppie di microtubuli saldati a formare un anello intorno a due microtubuli centrali. Si distinguono per la lunghezza e il numero in cui sono presenti sulla membrana plasmatica: le ciglia sono numerose e corte (10-25 µm), i flagelli sono pochi e lunghi (50-75 µm). I loro movimenti coordinati muovono la cellula nell’ambiente circostante o creano delle correnti nel liquido extracellulare che provocano un flusso costante delle particelle sospese.
La cellula vegetale
La cellula vegetale possiede alcune strutture esclusive, tra cui una parete cellulare e organuli quali i plastidi (comprendenti i cloroplasti) e i vacuoli. La parete cellulare è una struttura esterna alla membrana plasmatica ed è costituita per la massima parte da polisaccaridi, soprattutto cellulosa. Essa conferisce sostegno e forma alla cellula vegetale.

Struttura della cellula vegetale
I plasmodesmi sono giunzioni cellulari tipiche delle cellule vegetali: sono canali di citoplasma rivestiti di membrana che attraversano le pareti di due cellule vicine e permettono il passaggio diretto di alcune molecole.
I plastidi sono organuli avvolti da una doppia membrana, distinti in cloroplasti, cromoplasti e leucoplasti in base al loro colore.
I cloroplasti provvedono al rifornimento dell’energia necessaria alla cellula: ciò avviene per cattura dell’energia solare mediante il processo di fotosintesi, opera to da un pigmento verde, la clorofilla, presente in un complesso sistema di membrane (tilacoidi) contenuto nel citoplasma (stroma); le membrane dei tilacoidi sono impilate l’una sull’altra a formare delle pile (grani). L’energia solare catturata viene utilizzata da altre molecole per sintetizzare glucosio a partire da diossido di carbonio e acqua. Come i mitocondri, anche i cloroplasti contengono ribosomi e un proprio DNA.

Cellule vegetali al cui interno sono visibili i cloroplasti
I cromoplasti contengono il pigmento rosso-arancione carotene; sono presenti nei fiori e nei frutti e servono ad attirare gli insetti per consentire l’impollinazione.
I leucoplasti sono privi di pigmenti e perciò bianchi; si trovano per esempio nei tuberi, dove immagazzinano sostanze di riserva come l’amido, che deriva dalla trasformazione degli zuccheri prodotti dalla fotosintesi.
I vacuoli sono cavità nelle quali si accumulano acqua, prodotti di rifiuto o sostanze nutritive. Le cellule vegetali spesso presentano un unico, grosso vacuolo centrale ripieno d’acqua, che occupa fino al 90% dello spazio interno e che, premendo sulla membrana plasmatica, contribuisce a mantenere turgida la cellula.
Il differenziamento delle cellule negli organismi pluricellulari
Negli organismi unicellulari l’unica cellula deve svolgere da sola tutte le funzioni necessarie all’organismo. Perciò le modificazioni di forma sono transitorie (legate a situazioni ambientali) e reversibili. Soltanto negli organismi pluricellulari le varie cellule sono differenziate, in quanto assumono forme molto diverse, rispondenti a funzioni altamente specializzate.
Il differenziamento cellulare è un processo irreversibile: la cellula che si differenzia perde la possibilità di assumere altre specializzazioni e la capacità di compiere alcune funzioni proprie delle cellule in generale (per esempio, le cellule nervose non si riproducono). In alcuni casi possono scomparire certe strutture (per esempio, nel sangue le cellule che costituiscono i globuli rossi maturi sono prive di nucleo, mitocondri e ribosomi). Al tempo stesso le cellule differenziate acquistano funzioni e strutture nuove (i globuli rossi accumulano grandi quantità della proteina emoglobina, capace di trasportare ossigeno).

Dall'atomo all'organismo
Per il buon funzionamento dell’organismo pluricellulare diventa perciò indispensabile che le cellule interagiscano continuamente tra loro e svolgano azioni coordinate. La comunicazione tra cellule può avvenire tramite siti di contatto specializzati detti giunzioni comunicanti: queste possono essere, per esempio, le sinapsi chimiche tra le cellule nervose, i plasmodesmi tra le cellule vegetali, le giunzioni strette, tipiche dei tessuti epiteliali e le giunzioni gap, tipi che del tessuto cardiaco e formate da subunità di proteine che possono aprirsi o chiudersi per regolare il passaggio di piccole molecole. A tale scopo le cellule che si sono specializzate per svolgere la stessa funzione si associano formando i tessuti; a loro volta i tessuti si uniscono per costituire un organo in grado di svolgere una determinata funzione; più organi possono unirsi in un apparato o un sistema e cooperare per svolgere una funzione complessa: un apparato è un insieme di organi molto diversi tra loro per il tipo di tessuto che li costituisce. Per esempio, il naso e il polmone nel caso dell’apparato respiratorio. Un sistema, invece, è formato da organi costituiti dallo stesso tipo di tessuto. Per esempio, il cervello e i nervi sono fomati da tessuto nervoso, per cui si parla di sistema nervoso.
Ogni apparato e ogni sistema svolgono una funzione specifica, ma il loro lavoro è integrato per mantenere l’interno del corpo nelle condizioni necessarie alla vita e al funzionamento delle cellule e consentire così la sopravvivenza dell’organismo.
L’attività della cellula
La cellula è un “sistema aperto”, che preleva dall’ambiente esterno le sostanze nutritive e vi immette i prodotti di rifiuto. Il passaggio di materiali avviene attraverso la membrana plasmatica: essa partecipa attivamente alle reazioni cellulari regolando il trasporto di sostanze da e verso la cellula. La cellula deve prelevare dall’ambiente anche l’energia necessaria per attivare tutte le reazioni: questa può essere energia chimica, contenuta negli alimenti e liberata attraverso la respirazione cellulare e la fermentazione, oppure energia solare, utilizzata attraverso la fotosintesi. La fotosintesi è l’unico sistema a disposizione degli esseri viventi in grado di trasformare l’energia solare in energia chimica, direttamente utilizzabile dalle cellule per svolgere tutti i loro processi metabolici. Gli organismi eterotrofi devono procurarsi il glucosio (combustibile per la sintesi di ATP) nutrendosi degli organismi autotrofi (piante, alghe, alcuni batteri): questo rapporto di dipendenza è alla base delle catene alimentari della biosfera. Inoltre la fotosintesi libera come sottoprodotto l’ossigeno, utilizzato da tutti gli esseri viventi per la respirazione.
Il trasporto dei materiali
attraverso la membrana plasmatica Per soddisfare le sue esigenze biologiche di adattamento all’ambiente e di riproduzione, la cellula deve introdurre al suo interno i materiali che le servono per la nutrizione e la respirazione e, al tempo stesso, eliminare i prodotti di rifiuto (escrezione).
Il passaggio di materiali tra l’ambiente interno della cellula e l’ambiente extracellulare è regolato dalla membrana plasmatica. Il suo doppio strato lipidico costituisce una barriera invalicabile per la maggior parte delle grosse molecole biologiche – che sono polari e idrosolubili – e degli ioni, per consentire il passaggio delle grosse molecole e degli ioni necessari alla vita della cellula, intervengono le proteine della membrana, alcune delle quali formano pori o canali, mentre altre sono dotate di siti di legame per specifiche moleco-
le. Per le sue caratteristiche selettive riguardo il passaggio di molecole e ioni, la membrana plasmatica viene detta semipermeabile. Il passaggio delle sostanze attraverso la membrana può avvenire secondo due meccanismi: il primo non implica consumo di energia e viene detto trasporto passivo; il secondo comporta consumo di energia da parte della cellula e si distingue in trasporto attivo, endocitosi ed esocitosi.

Alcune delle possibilità di movimento di molecole tra citoplasma (1) e spazio extracellulare (2). Trasporto passivo: A. Diffusione; B. Diffusione facilitata; Trasporto attivo:
_D. Trasporto primario (contro gradiente); C.-E. Trasporto secondario: C. Uniporto; E. Simporto; Esocitosi/Endocitosi: F.-G. Esocitosi” >}}
Il trasporto passivo
Il trasporto passivo è il movimento di sostanze in soluzione attraverso la membrana che non richiede dispendio di energia da parte della cellula. La membrana non interferisce sulla direzione del movimento delle particelle, poiché questo avviene grazie all’esistenza di gradienti, cioè di differenze di concentrazione: le particelle si muovono spontaneamente da zone ad alta concentrazione a zone a bassa concentrazione, quali quelle esistenti rispettivamente all’esterno e all’interno della membrana stessa (il movimento
procede fino all’instaurarsi di condizioni di equilibrio). In altre parole, il movimento delle particelle avviene sfruttando l’energia potenziale dovuta alla presenza del gradiente di concentrazione.
Esistono tre forme di trasporto passivo: la diffusione semplice, la diffusione facilitata e l’osmosi.
La diffusione semplice è il movimento di gas o molecole liposolubili attraverso il doppio strato fosfolipidico della membrana plasmatica. La velocità di diffusione è influenzata dalla concentrazione della sostanza che diffonde, dalla temperatura e dalla pressione. La diffusione prosegue finché non viene annullato il gradiente di concentrazione,
quando cioè la concentrazione del gas o della molecola diventa uguale ai due lati della membrana.
La diffusione facilitata è il trasporto di ioni, amminoacidi e monosaccaridi mediato da proteine della membrana. Alcune di queste (proteine canale) possono formare dei canali permanenti attraverso la membrana. Altre proteine (trasportatori) possiedono dei siti di legame specifici per determinate molecole; una volta avvenuto il legame, il trasportatore modifica la propria forma e, di conseguenza, la molecola legata viene trasferita e liberata dall’altro lato della membrana. Poiché la membrana plasmatica ha un numero limitato di trasportatori, la velocità della diffusione facilitata è minore di quella della diffusione semplice.
L’osmosi è la diffusione dell’acqua attraverso la membrana plasmatica. Il passaggio dell’acqua è consentito dalla presenza di pori formati dall’assemblaggio di un certo numero
di proteine intrinseche. L’osmosi termina quando viene raggiunto l’equilibrio osmotico: in queste condizioni, la velocità del flusso d’acqua ai due lati della membrana plasmatica è uguale nei due sensi e non vi è più un passaggio netto di molecole da un lato all’altro.
Le tre forme di trasporto passivo, dipendendo da differenze (gradienti) di concentrazione, sono reversibili, cioè avvengono in un senso o nell’altro a seconda del gradiente.
Il trasporto attivo
Il trasporto attivo è il movimento di materiali attraverso la membrana plasmatica contro il gradiente di concentrazione (cioè da zone a bassa concentrazione a zone ad alta concentrazione) e con consumo di energia da parte della cellula. Le sostanze trasportate dall’interno all’esterno possono essere prodotti di rifiuto (la cui presenza, anche in quantità piccolissime, potrebbe danneggiare la cellula);
quelle trasportate dall’esterno all’interno sono sostanze nutritive presenti in bassa concentrazione nell’ambiente extracellulare e ioni che devono essere presenti nella cellula a concentrazioni inferiori rispetto a quelle del liquido extracellulare.
Il trasporto attivo è regolato da proteine intrinseche della membrana (chiamate pompe perché spostano molecole contro il gradiente di concentrazione), provviste di siti di legame per una molecola specifica da trasportare e per l’ATP. L’idrolisi (scissione) dell’ATP fornisce
l’energia necessaria alla proteina trasportatrice per cambiare forma e trasportare la molecola attraverso la membrana. Una volta liberata la molecola, la proteina trasportatrice riacquista la configurazione originaria.
L’endocitosi e l’esocitosi sono due forme di trasporto attivo alle quali la cellula ricorre se le sostanze che deve introdurre o eliminare sono interi batteri o macromolecole molto grandi.
L‘endocitosi è il trasporto di materiali all’interno della cellula per mezzo di vescicole. La cellula circonda la particella da introdurre con la sua membrana plasmatica, fino a inglobarla in una vescicola che poi si fonde con un lisosoma; all’interno di quest’ultimo la particella assunta
viene demolita dagli enzimi proteolitici. L’endocitosi rappresenta il metodo di assunzione degli alimenti di alcuni protozoi come l’ameba. In questo caso il processo prende il nome di fagocitosi: l’ameba circonda l’alimento con prolungamenti del citoplasma detti pseudopodi, quindi lo ingloba e lo digerisce servendosi dei propri lisosomi. Anche i globuli bianchi del sangue ricorrono alla fagocitosi per inglobare e distruggere batteri e altri corpi estranei.
L‘esocitosi è il trasporto di materiali al di fuori della cellula. In pratica, è l’opposto dell’endocitosi: le sostanze che devono essere rimosse dalla cellula vengono racchiuse in un vacuolo, che si sposta fino alla periferia della cellula in modo da fondersi con la membrana plasmatica.
Le vie metaboliche
Il metabolismo è l’insieme delle reazioni chimiche che avvengono nella cellula. Nel metabolismo si distinguono due fasi: il catabolismo (comprendente tutte le reazioni di demolizione delle molecole fornite dall’ambiente sotto forma di sostanze nutritive) e l’anabolismo (comprendente tutte le reazioni di sintesi delle macromolecole biologiche e il lavoro cellulare come il movimento, la divisione, il trasporto attraverso la membrana).
Le reazioni cataboliche sono reazioni esoergoniche in quanto avvengono con liberazione di parte dell’energia contenuta nei legami chimici delle molecole demolite. Al contrario, le reazioni anaboliche sono reazioni endoergoniche, cioè avvengono con consumo di energia. L’energia necessaria per tutte le reazioni deriva dall’energia chimica dei legami presenti nelle molecole delle sostanze alimentari o dall’energia radiante proveniente dal Sole.
Le reazioni del metabolismo sono spesso raggruppate in vie metaboliche. Per via metabolica si intende una sequenza di reazioni controllate da enzimi, concatenate in modo tale che i prodotti di alcune di tali reazioni fungono da reagenti per altre. Le vie metaboliche principali sono la fotosintesi, attraverso la quale l’energia solare viene catturata e immagazzinata come energia chimica, e la glicolisi, completata dalla respirazione cellulare o dalla fermentazione, attraverso cui l’energia chimica viene estratta e resa disponibile alle cellule. La fotosintesi è svolta dagli organismi autotrofi (le piante, le alghe e alcuni batteri), che sono cioè in grado di produrre le proprie sostanze alimentari a partire da sostanze inorganiche semplici. Gli organismi eterotrofi, invece, per il loro fabbisogno alimentare dipendono dai materiali organici elaborati e sintetizzati dagli organismi autotrofi (sono organismi eterotrofi i funghi, gli animali e la maggior parte dei batteri).
Esistono altre vie metaboliche, sfruttate soprattutto dai batteri, che utilizzano come fonti d’energia il metano (metanobatteri), composti dello zolfo (solfobatteri), dell’azoto (batteri nitrificanti) e del ferro (ferrobatteri). Tra i batteri nitrificanti ricordiamo i nitrosobatteri (per esempio, il Nitrosomonas), che trasformano lo ione ammonio (NH4+) in nitrito (NO2-), e i nitrobatteri (per esempio, il Nitrobacter), che trasformano il nitrito in nitrato (NO3-), direttamente utilizzabile dalle piante che lo incorporano in molecole biologiche.
La fotosintesi
La fotosintesi è il processo attuato dagli organismi autotrofi per produrre glucosio a partire da acqua e diossido di carbonio (anidride carbonica), utilizzando come fonte di energia la luce solare assorbita da un particolare pigmento fotosensibile, la clorofilla.
La fotosintesi si svolge all’interno dei cloroplasti, nei quali si trovano due varietà del pigmento verde clorofilla, la clorofilla a e la clorofilla b, sensibili a due lunghezze d’onda leggermente diverse. Entrambi i tipi di clorofilla sono in grado di assorbire la luce blu e quella rossa, mentre non assorbono la luce verde e quella gialla, che vengono riflesse (è per questo che le cellule contenenti i cloroplasti appaiono di colore verde).
La reazione complessiva della fotosintesi è schematizzabile nella formazione di 1 molecola di glucosio (con legami ricchi di energia) e di 6 molecole di ossigeno a partire da 6 molecole di diossido di carbonio e 6 molecole di acqua:
_6CO2 + 6H2O ⟶ C6H12O6 + 6O2” >}}
_Reazione complessiva della fotosintesi: l’anidride carbonica,a assieme all’acqua, viene trasformata in glucosio e ossigeno” >}}
Il meccanismo attraverso il quale si compie la fotosintesi può essere scisso in due fasi distinte (fase luminosa e fase oscura), ciascuna costituita da una complessa serie di reazioni catalizzate da enzimi. Queste fasi sono collegate tra loro e avvengono in due punti diversi del cloroplasto.
Reazioni della fase luminosa
Nella fase luminosa la luce assorbita dalla clorofilla viene utilizzata come fonte di energia per rompere le molecole di acqua e per sintetizzare ATP (da ADP) e NADPH (da NADP+).

Le reazioni della fase luce dipendente della fotosintesi clorofilliana.
La clorofilla, alcuni pigmenti accessori e molecole trasportatrici di elettroni sono disposti sulle membrane dei tilacoidi a formare due raggruppamenti, il fotosistema I e il fotosistema II. L’energia luminosa “catturata” dai pigmenti del fotosistema II viene trasferita a una particolare molecola di clorofilla, che costituisce il centro reattivo. La clorofilla-centro reattivo viene “eccitata” e perde un suo elettrone che entra nella catena dei trasportatori di elettroni. Il flusso dell’elettrone lungo la catena fornisce l’energia necessaria per la sintesi di ATP da ADP.
Anche il fotosistema I viene raggiunto dall’energia solare e la clorofilla-centro reattivo perde un elettrone; questo va alla catena dei trasportatori di elettroni del fotosistema I, il cui ultimo accettore è una molecola di NADP+ (nicotinammide-adenin-dinucleotide fosfato). Ciascuna molecola di NADP+ si lega a due elettroni e a uno ione idrogeno (proveniente dalla dissociazione dell’acqua), formando NADPH, un trasportatore ricco di energia. In seguito alla dissociazione dell’acqua, si libera ossigeno.
Reazioni della fase oscura
Nella fase oscura, gli enzimi presenti nello stroma utilizzano l’energia chimica contenuta nell’ATP e nel NADPH per ridurre (o fissare) il diossido di carbonio a glucosio. Le reazioni di questa fase, che possono avvenire anche in assenza di luce, costituiscono il ciclo di Calvin-Benson, dal nome degli scopritori.

Schema riassuntivo delle reazioni della fase luminosa e della fase oscura della fotosintesi
Durante il ciclo di Calvin-Benson, una molecola di ribulosio difosfato (RuBP), uno zucchero a 5 atomi di carbonio già presente nel cloroplasto grazie a precedenti reazioni, si lega con una molecola di diossido di carbonio, CO2 , formando un composto a 6 atomi di carbonio molto instabile. Quest’ultimo reagisce con una molecola d’acqua, producendo due molecole di acido fosfoglicerico (PGA), un composto a 3 atomi di carbonio cui è legato un gruppo fosfato. Successivamente, il PGA viene trasformato in fosfogliceraldeide (PGAL) nel corso di una reazione che richiede NADPH e ATP provenienti dalla fase luminosa.
![alt_text]https://upload.wikimedia.org/wikipedia/commons/thumb/2/27/Calvin-cycle3.png/699px-Calvin-cycle3.png" title=“Ciclo di Calvin” >}}
{kind=link}
Iniziando il ciclo di Calvin-Benson con 6 molecole di RuBP, si ottengono 12 molecole di PGAL: di queste, 10 sono utilizzate per rigenerare il RuBP che verrà impiegato in un nuovo ciclo, mentre 2 si combinano a formare una molecola di glucosio. Alla fine del ciclo i trasportatori ATP e NADPH si sono “esauriti”, trasformati rispettivamente in ADP e NADP+, e tornano alle reazioni della fase luminosa per essere nuovamente “caricati”.
Il glucosio può essere demolito nel corso della respirazione cellulare, oppure può essere convertito in lipidi o altri costituenti della cellula, o, ancora, immagazzinato sotto forma di amido o trasformato in cellulosa.
La liberazione dell’energia
Tutti gli organismi, sia autotrofi sia eterotrofi, ottengono l’energia necessaria al proprio metabolismo liberando l’energia chimica contenuta nei legami delle sostanze alimentari e trasformandola nei legami altamente energetici dell’ATP. La successiva demolizione dell’ATP porta alla liberazione graduale di energia, che la cellula può utilizzare per attivare le sue reazioni. La fonte primaria di energia per la cellula è quella accumulata come energia chimica nei legami del glucosio.
La glicolisi è l’insieme delle reazioni che demoliscono una molecola di glucosio fino a formare 2 molecole di acido piruvico, un composto a 3 atomi di carbonio. La glicolisi avviene nel citoplasma della cellula senza richiedere l’intervento di ossigeno: utilizzando l’energia di 2 molecole di ATP viene liberata energia sufficiente a trasformare 4 molecole di ADP in altrettante di ATP. Inoltre si liberano 4 ioni idrogeno che si legano a 2 molecole di NAD+ per formare NADH (un trasportatore di elettroni ricco di energia, che verrà utilizzato nelle reazioni successive). In sintesi durante la glicolisi:
_C6H12O6 + 2ATP ⟶ 2CH3COCOOH + 4ATP + 2NADH” >}}
_(in termini di ATP il bilancio netto è di 2ATP prodotti)” >}}
A questo punto l’acido piruvico può essere ulteriormente demolito in presenza di ossigeno mediante la respirazione cellulare o, in assenza di ossigeno, con la fermentazione.
La respirazione cellulare
La respirazione cellulare è un processo aerobico che comporta la formazione di ATP e la completa degradazione dell’acido piruvico fino ad acqua e diossido di carbonio. Avviene nei mitocondri e comprende 3 sistemi interdipendenti di reazioni: 1. trasformazione dell’acido piruvico in acetil-CoA; 2. ciclo di Krebs; 3. sistema di trasporto degli elettroni.

Schema della respirazione cellulare
- L’acido piruvico entra nei mitocondri, dove si trasforma in acetaldeide, un composto a 2 atomi di carbonio; il carbonio rimosso viene liberato sotto forma di diossido di carbonio (eliminato dalla cellula come prodotto di rifiuto) mentre gli ioni idrogeno si legano a molecole di NAD + formando NADH. L’acetaldeide si lega al coenzima A (CoA) costituendo il complesso acetil-CoA: in pratica, da 2 molecole di acido piruvico si formano 2 acetil-CoA.
- L’acetil-CoA entra nel ciclo di Krebs, o ciclo dell’acido citrico, nel corso del quale vengono liberati 2 atomi di carbonio sotto forma di diossido di carbonio, viene prodotta 1 molecola di ATP, 1 coppia di ioni idrogeno si lega al trasportatore FAD (flavin-adenin-dinucleotide), che diventa FADH 2 , mentre altre 3 si legano al NAD + , che diventa NADH. Nel corso delle reazioni alle quali prende parte il coenzima A non subisce modificazioni, per cui al termine del ciclo di Krebs viene liberato e reso disponibile per legarsi a un’altra molecola di acetaldeide.
- Il NADH e il FADH2 entrano nella catena di trasporto degli elettroni, costituita da una serie di proteine (citocromi), ordinate sulle creste dei mitocondri. Gli elettroni degli
atomi di idrogeno vengono “trasferiti” da un trasportatore all’altro cedendo progressivamente la loro energia, che verrà utilizzata per “pompare” ioni idrogeno nello spazio interno del mitocondrio. Il flusso di ioni idrogeno fornisce l’energia necessaria alla sintesi di ATP.
Il ciclo di Krebs
Il ciclo di Krebs consiste in una serie di reazioni in cui sono convogliati i prodotti della glicolisi, cioè della demolizione delle sostanze alimentari (zuccheri, grassi, proteine) per essere ulteriormente degradati fino ad acqua (H2O) e diossido di carbonio (CO2), rendendo così disponibile per la cellula l’energia chimica dei loro legami (sotto forma di ATP).

Schema del Ciclo di Krebs
Tali prodotti, trasformati in acido piruvico, sono immessi nel ciclo sotto forma di acetil-coenzima A (o acetil-CoA), molecola ottenuta dall’unione di un gruppo acetilico (a 2 atomi di carbonio) con il coenzima A, presente all’interno dei mitocondri. Nella prima delle nove reazioni di cui si compone il ciclo, l’acetil-CoA reagisce con l’ossalacetato, formandoun composto a 6 atomi di carbonio: l’acido citrico. Questo viene ossidato nel corso delle reazioni successive formando diversi composti intermedi, fino alla riformazione della molecola di ossalacetato che “chiude” un giro del ciclo.
Le ossidazioni avvengono attraverso l’azione di coenzimi, detti anche trasportatori di elettroni, che vengono ridotti, ossia assumono gli elettroni carichi di energia provenienti dalla rottura dei legami. I coenzimi coinvolti sono NAD (nicotinammide-adenin-dinucleotide) e FAD (flavin-adenin-dinu-cleotide) i quali, riducendosi, diventano rispettivamente NADH e FADH.
Il bilancio energetico del ciclo di Krebs è di una sola molecola di ATP, essendo la maggior parte dell’energia contenuta nelle molecole dei trasportatori di elettroni, che la renderanno disponibile per la cellula dopo essere stati ossidati a opera degli enzimi della cosiddetta catena respiratoria. Alla fine della catena respiratoria gli elettroni, insieme ad altrettanti ioni idrogeno, si combinano con l’ossigeno per formare acqua. Il rendimento energetico complessivo della respirazione cellulare a partire da 1 molecola di glucosio è di 38 molecole di ATP:
- 2 sono prodotte nella glicolisi;
- 2 nel ciclo di Krebs;
- 34 nel sistema di trasporto degli elettroni.
In un certo senso si può considerare la respirazione cellulare l’inverso della fotosintesi: i reagenti della respirazione cellulare sono infatti i prodotti finali della fotosintesi. Inoltre, mentre la fotosintesi è un processo endoergonico, la respirazione cellulare è complessivamente esoergonica.
| FOTOSINTESI | RESPIRAZIONE CELLULARE | |
| reagenti | CO2 e H2O | C6H12O6 |
| progetti | C6H12O6 e O2 | CO2 e H2O |
| energia | è assorbita | è liberata |
| equazione complessiva | 6CO2+6H2O⟶ C6H1206 + 6O2 | C6H12O6 + 6O2 ⟶ 6CO2 + 6H2O |
_Confronto tra fotosintesi e respirazione cellulare” >}}
La fermentazione
La fermentazione è un complesso di reazioni anaerobiche nel corso delle quali l’acido piruvico viene trasformato o in alcol etilico (o etanolo) e diossido di carbonio (fermentazione alcolica) o in acido lattico (fermentazione lattica). In entrambi i casi vengono impiegati atomi di idrogeno ed elettroni del NADH, che ritorna NAD+, nuovamente utilizzabile per la glicolisi.
La fermentazione è il modo con cui alcuni microrganismi (funghi e batteri) utilizzano per le proprie necessità energetiche sostanze organiche (zuccheri, ma anche proteine, acidi e alcoli). Ha un rendimento energetico molto inferiore alla respirazione cellulare. Può avvenire anche nei tessuti animali: per esempio, la fermentazione lattica si verifica nei muscoli, quando a causa di un’attività fisica troppo intensa o prolungata la disponibilità di ossigeno è insufficiente allo svolgimento della respirazione cellulare.
Il linguaggio della cellula
Gli acidi nucleici, DNA (acido desossiribonucleico) e RNA (acido ribonucleico) sono composti di straordinaria importanza biologica, essendo responsabili dell’informazione genetica, che trasmettono attraverso il processo riproduttivo di generazione in generazione, e del controllo della sintesi proteica nelle cellule. Il “programma” di istruzioni è “scritto” in forma molecolare nella sequenza dei nucleotidi che costituiscono le unità costruttive del DNA (codice genetico). Poiché il DNA è confinato nel nucleo della cellula, le sue istruzioni vengono copiate su catene di un tipo di RNA (RNA messaggero), che provvede a trasportarle nei siti dove avviene la sintesi proteica attraverso due altri tipi di RNA (RNA di trasporto e RNA ribosomiale).
La struttura del DNA e dell’RNA
Il DNA e l’RNA sono due tipi di acidi nucleici formati da subunità chiamate nucleotidi: sono pertanto dei polinucleotidi (il DNA è formato da due catene o filamenti di polinucleotidi avvolte a doppia elica; l’RNA è fomato da una singola catena polinucleotidica). Ogni nucleotide è costituito da tre componenti:
- un gruppo fosfato;
- uno zucchero a 5 atomi di carbonio;
- una base azotata.

Elementi costituitivi dei nucleotidi
Nel DNA lo zucchero è il desossiribosio, che può essere legato a quattro basi azotate differenti: adenina (A); timina (T); guanina (G); citosina ©. Nell’RNA lo zucchero è il ribosio e, come nel DNA, può essere legato a quattro basi azotate differenti: tre sono comuni al DNA (adenina, guanina e citosina); la quarta è differente e prende il nome di uracile (U).
Delle 5 basi azotate, l’adenina e la guanina sono dette puriniche, in quanto derivano dalla purina, un composto eterociclico azotato (formato da un anello pentaatomico condensato con un anello esaatomico). La citosina, la timina e l’uracile sono dette pirimidiniche, derivando dalla pirimidina, un altro composto eterociclico azotato (formato da un anello esaatomico). Il gruppo fosfato di ogni nucleotide si lega allo zucchero del nucleotide seguente per mezzo di un legame covalente e in tal modo si forma una lunga catena che può essere paragonata a un pettine: i “denti” che sporgono corrispondono alle basi azotate, che sono legate a un’impalcatura formata da molecole di zucchero alternate a gruppi fosfato.
Il DNA è presente nel nucleo di tutte le cellule, di cui porta il codice genetico. L’RNA si trova sia nel nucleo sia nel citoplasma delle cellule e partecipa direttamente alla sintesi delle proteine.

Struttura a doppia elica del DNA
La molecola del DNA
Grazie agli studi compiuti nel 1953 da due scienziati, l’inglese Francis Crick (1916) e l’americano James Watson (1928), sappiamo che la molecola del DNA è formata da due catene orientate in direzioni opposte: in una i nucleotidi sono disposti nella sequenza 3’-5’ (i numeri si riferiscono agli atomi di carbonio del desossiribosio), nell’altra nella sequenza 5’-3’. Le due catene sono avvolte su se stesse in modo da formare una doppia elica, paragonabile a una scala a chiocciola: le due “ringhiere” sono date dall’alternanza dello zucchero e del gruppo fosfato, mentre i “gradini” dalle coppie di basi azotate, unite tra loro da deboli legami a idrogeno. Per la regola dell’appaiamento delle basi, una base purinica può appaiarsi solo con una base pirimidinica: più precisamente, l’adenina (A) può accoppiarsi solo con la timina (T) e la guanina (G) solo con la citosina ©. In questo modo tutti i “gradini” sono della stessa lunghezza e le due “ringhiere” restano perfettamente parallele. L’alternanza del desossiribosio e del gruppo fosfato è uguale in tutte le specie viventi; è invece variabile e caratteristico l’ordine con cui si dispongono le due coppie di basi azotate (A-T e C-G).

Struttura chimica del DNA. Le sequenze possono essere definite senso e non senso.
I tre tipi di RNA
Esistono tre tipi diversi di RNA:
- RNA messaggero, o m-RNA
- RNA di trasporto, o t-RNA
- RNA ribosomiale, o r-RNA
I tre tipi di RNA sono preposti ognuno a una funzione specifica nella sintesi delle proteine. Le molecole di RNA hanno dimensioni e peso molecolare differenti, ma la struttura si mantiene costante.
L‘RNA messaggero è una copia a filamento singolo complementare di un tratto di DNA e “trascrive” le istruzioni per la corretta sequenza nella quale gli amminoacidi devono unirsi tra loro per formare una proteina. Ogni gruppo di 3 nucleotidi in sequenza sull’m-RNA è detto codone e porta le informazioni necessarie alla sintesi di un particolare amminoacido.
L’RNA di trasporto (t-RNA) riconosce le informazioni dell’m-RNA e interviene nella sintesi degli amminoacidi. In alcuni tratti dell’RNA di trasporto le basi si appaiano per complementarietà. Si ha così un avvicendarsi di tratti appaiati e di “occhielli” a basi disaccoppiate che conferiscono alla molecola una particolare forma a “trifoglio”. Un’estremità del t-RNA si lega a uno specifico amminoacido; dalla parte opposta, una particolare sequenza di 3 basi azotate (anticodone) si appaia a un codone di m-RNA.
L’RNA ribosomiale si lega ad alcune proteine per formare i ribosomi, organuli composti da due subunità di dimensioni diverse sui quali avviene la sintesi proteica.
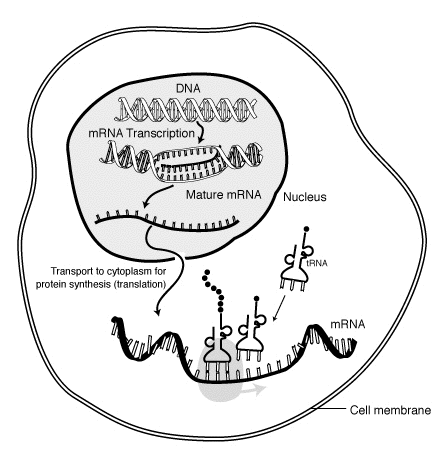
Il "ciclo vitale" di un mRNA in una cellula eucariote. L'RNA è trascritto nel nucleo cellulare; dopo essere stato completamente modificato viene trasportato nel citoplasma e tradotto da un ribosoma. Alla fine della sua vita l'mRNA viene degradato.
| TIPO | N. DI TIPI DELLA CELLULA | FUNZIONE |
| m-RNA | 1000-10000 | trascrizione dell'informazione contenuta nel DNA e suo trasferimento ai ribosomi durante la sintesi proteica |
| t-RNA | 50-60 | trasporto di amminoacidi e riconoscimento dei codoni dell'm-RNA durante la sintesi proteica |
| r-RNA | 2 | formazione di ribosomi (in associazione con proteine specifiche) |
_I principali tipi di RNA” >}}
Il codice genetico
Il codice genetico è il sistema per cui le informazioni genetiche codificate nel DNA arrivano a operare la sintesi di tutte le proteine necessarie alla vita degli organismi. Il suo linguaggio si basa su un “alfabeto” molecolare rappresentato dalla sequenza dei nucleotidi del DNA, che viene tradotto nella sequenza degli amminoacidi di una proteina. Il codice genetico (v. fig. 5.5) dispone di 4 “lettere” (le 4 diverse basi azotate) per specificare i 20 amminoacidi. Utilizzando gruppi di 3 nucleotidi (triplette, o codoni) si ottengono 43 = 64 combinazioni diverse. Tre di queste triplette (triplette non senso) non corrispondono a nessun amminoacido: esse servono per segnalare la fine della catena proteica. La tripletta AUG indica l’inizio della catena proteica; a essa corrisponde anche l’amminoacido metionina. Si definisce gene la sequenza di triplette che codifica una proteina.
Tabella codifica codoni nel DNA
Il codice genetico è ridondante, poiché uno stesso amminoacido è codificato da più di una tripletta. Le triplette che codificano lo stesso amminoacido sono molto simili e generalmente differiscono solo per l’ultima delle tre basi. Ciò ha suggerito l’ipotesi che l’informazione fondamentale sia contenuta nelle prime due basi e che la terza serva a garantire una maggiore precisione.
Il codice genetico è universale, dal momento che è identico in tutti gli esseri viventi (ogni tripletta ha lo stesso significato per tutti gli organismi).
La sintesi proteica
Si definisce sintesi proteica il processo con cui una sequenza di nucleotidi viene convertita nella successione di amminoacidi formanti una proteina. Alla sintesi proteica prendono parte attiva l’m-RNA, il t-RNA e l’r-RNA.
L’m-RNA copia l’informazione contenuta nel DNA e la trasporta dal nucleo al citoplasma (questo stadio è detto trascrizione); il t-RNA e l’r-RNA traducono il messaggio scritto sull’m-RNA in una sequenza di amminoacidi (questo stadio è detto traduzione). Durante la sintesi proteica perciò, l’informazione genetica passa dal DNA all’RNA e dall’RNA alle proteine. È questo il dogma centrale della biologia.
La trascrizione
La trascrizione è lo stadio della sintesi proteica in cui le informazioni sono trasferite dal DNA all’RNA, secondo le regole dell’appaiamento delle basi complementari. Come nella replicazione, è necessario che le basi azotate sporgano dalla doppia elica del DNA. Perciò il tratto di DNA che deve essere trascritto viene aperto in un punto ben preciso, caratterizzato dalla tripletta AUG di “inizio lettura”. Un enzima, l’RNA-polimerasi, si lega a uno dei due filamenti di DNA che serve da “stampo”, e procede dall’estremità 3’ all’estremità 5’ legando i ribonucleotidi complementari presenti nel nucleo. Si forma in questo modo l’m-RNA.
Quando l’RNA-polimerasi giunge alla tripletta di “fine lettura”, l’m-RNA si separa dalla catena di DNA. Negli eucarioti vi è una quantità di DNA molto superiore a quella necessaria per la sintesi delle proteine. Infatti, i geni degli eucarioti sono costituiti da sequenze di nucleotidi che codificano una proteina (esoni) alternati a sequenze che non codificano nulla (introni). Quando un gene così strutturato viene trascritto, si forma dapprima un lungo filamento di m-RNA (trascritto primario) che comprende sia gli esoni sia gli introni. Successivamente gli introni vengono rimossi e le estremità degli esoni saldate insieme (processamento dell’mRNA). Solo a questo punto il filamento di m-RNA, detto maturo , passa per i pori della membrana nucleare ed entra nel citoplasma, dove si lega ai ribosomi. Il DNA “modello” si riavvolge a formare la doppia elica, oppure si lega a una nuova molecola di RNA-polimerasi per sintetizzare un nuovo filamento di m-RNA.
La traduzione
La traduzione è lo stadio della sintesi proteica in cui le istruzioni portate dall’m-RNA vengono tradotte nella sequenza corretta di amminoacidi per formare una proteina.
La traduzione ha luogo nel ribosoma (formato da r-RNA e proteine), composto da due subunità: quella piccola contiene un sito di legame per l’m-RNA; quella grande ha due siti di legame per due molecole di t-RNA e un sito che catalizza la formazione del legame peptidico tra due amminoacidi adiacenti.
Ogni molecola di t-RNA è specifica per un unico amminoacido ed è in grado di riconoscere sia l’amminoacido che deve trasportare, sia il codone complementare di m-RNA associato al ribosoma.
La traduzione ha inizio quando due codoni del filamento di m-RNA si legano alla subunità piccola di un ribosoma. Il primo codone è la tripletta di “inizio lettura” AUG, alla quale corrisponde l’amminoacido metionina; il secondo codifica il primo vero amminoacido della proteina. I due t-RNA, che hanno rispettivamente l’anticodone di inizio e l’anticodone complementare al secondo codone, si legano alla subunità grande e si forma un legame peptidico (cioè il legame tra amminoacidi che forma le proteine) tra i due amminoacidi trasportati.
Il t-RNA di inizio si stacca dal ribosoma mentre il dipeptide (i due amminoacidi uniti dal legame peptidico) rimane legato al secondo t-RNA. Il ribosoma si sposta sopra un altro codone dell’m-RNA e una nuova molecola di t-RNA con il proprio amminoacido si dispone nel sito di legame vuoto del ribosoma. Si crea un nuovo legame peptidico e il tripeptide si salda
all’ultimo t-RNA. Il processo di allungamento della catena polipeptidica prosegue in questo modo finché tutte le triplette sono state tradotte e viene raggiunto il codone di “fine lettura”. La proteina completa si stacca dal ribosoma e specifici enzimi scindono il legame con la metionina.
Rappresentazione della traduzione e sintesi proteica nel ribosoma
La replicazione del DNA
Si definisce replicazione il processo di duplicazione semiconservativa del DNA. Il processo è definito semiconservativo poiché le due nuove doppie eliche di DNA sono formate entrambe da uno dei vecchi filamenti e da un nuovo filamento complementare. Grazie alla replicazione, la cellula che si sta dividendo raddoppia il proprio materiale genetico per trasmetterne una copia a ognuna delle due cellule figlie.
La replicazione prende avvio quando, in un punto preciso di inizio, l’enzima DNA-elicasi rompe i legami a idrogeno tra le basi azotate e un breve tratto della doppia elica di DNA si despiralizza. In questo modo sporgono le basi azotate del DNA originario, che servono da “modello” per la formazione del nuovo DNA.
Un altro enzima, la DNA-polimerasi, si sposta lungo ciascun filamento di DNA, dall’estremità 3’ all’estremità 5’, per riconoscere le basi esposte del filamento “modello” e legare a esse i nucleotidi liberi (precedentemente sintetizzati nel citoplasma e portati all’interno del nucleo) con le basi complementari. La DNA-polimerasi lega anche il gruppo fosfato di un nucleotide al desossiribosio del nucleotide seguente.
Si forma così un nuovo filamento di DNA complementare al DNA che fa da “stampo”.

La replicazione del DNA
Nelle cellule eucarioti la doppia elica di DNA si lega a particolari proteine, gli istoni, per formare fibre di cromatina. Durante la divisione cellulare, la cromatina si avvolge su se stessa dando origine a masserelle molto compatte, i cromosomi. Nelle cellule procarioti le due estremità della catena di DNA si congiungono e si forma un unico filamento circolare.
Nelle cellule procarioti, in cui il DNA è circolare, la replicazione inizia in un solo punto e procede nelle due direzioni opposte, finché non è stato replicato tutto l’anello. Nelle cellule eucarioti il processo avviene simultaneamente in diverse unità di replicazione, una dopo l’altra lungo tutta la doppia elica di DNA; al termine, tutte le unità saranno congiunte.
Per il corretto funzionamento delle cellule figlie è importante che la replicazione avvenga con il minor numero possibile di errori: l’enzima DNA-polimerasi ha la funzione di impedire o rimuovere appaiamenti sbagliati di nucleotidi. Nonostante ciò, durante la replicazione si possono verificare errori, anche se con una frequenza molto bassa (un nucleotide sbagliato su 100 milioni appaiati correttamente).
Questi errori provocano un cambiamento nella sequenza delle basi azotate, e di conseguenza dell’informazione genetica, e sono detti mutazioni. Le mutazioni sono alla base della variabilità genetica.
La riproduzione della cellula
Una delle caratteristiche fondamentali della cellula è la capacità di riprodursi. Gli organismi unicellulari si riproducono per divisione dell’unica cellula che li compone. Perciò in essi la riproduzione cellulare ha solo il compito di assicurare la continuità della loro specie nel tempo. Negli organismi pluricellulari la riproduzione assicura anche la crescita dell’individuo e la riparazione di parti danneggiate o usurate. Ricorrendo alla meiosi e alla riproduzione sessuale, gli organismi pluricellulari rimescolano il materiale genetico dei genitori per aumentare la variabilità genetica.
Il ciclo cellulare
Una cellula si divide quando si crea una sproporzione tra superficie e volume. Quando una cellula si accresce, il volume aumenta molto più della superficie, cioè della membrana plasmatica. In un tempo relativamente breve, i costituenti interni di una cellula in crescita avranno quindi a disposizione una superficie proporzionalmente meno estesa attraverso cui ottenere le sostanze nutritive ed eliminare i prodotti di rifiuto. Ciò compromette gravemente la vita della cellula, che di conseguenza deve dividersi.
Si definisce ciclo cellulare la sequenza di eventi compresa tra due divisioni cellulari successive. Negli organismi unicellulari (per esempio, batteri, amebe) il ciclo cellulare coincide con il ciclo vitale (cioè con l’insieme degli eventi tra una generazione e quella succesiva). Negli organismi pluricellulari il ciclo cellulare è in genere una parte del ciclo vitale (quello relativo alla crescita corporea, che si compie appunto attraverso ripetute divisioni cellulari). La maggior parte degli organismi pluricellulari completa infatti il proprio ciclo vitale attraverso la riproduzione sessuale, che comporta la fecondazione di una cellula uovo da parte di uno spermatozoo. Cellule uovo e spermatozoi sono cellule sessuali che si originano attraverso una particolare forma di di-visione cellulare chiamata meiosi.
I cromosomi
Al termine di un ciclo cellulare si formano due cellule figlie identiche alla cellula madre dalla quale derivano. Questa identità è garantita dalla duplicazione dell’informazione genetica completa della cellula madre nei nuclei delle due nuove cellule. Il materiale genetico è rappresentato dal DNA, che normalmente è disperso in tutto il nucleo come una massa confusa. Durante la divisione cellulare, il DNA si spiralizza formando i cromosomi (“corpi colorati”), costituiti da due bracci (cromatidi) uniti tra loro in corrispondenza di una regione particolare (centromero).

Disegno di un cromosoma eucariote duplicato e in metafase (1) Cromatidio – una delle due parti identiche dopo la fase G2. (2) Centromero – il punto di contatto dei due cromatidi e dove si legano i microtubuli. (3) Braccio corto. (4) Braccio lungo
Nella maggior parte delle cellule (cellule somatiche) si trovano coppie di cromosomi (ciascun membro della coppia è detto cromosoma omologo) con la stessa forma, la stessa lunghezza e un’informazione genetica molto simile: uno di origine paterna e uno di origine materna. Si definisce diploide la cellula che possiede coppie di cromosomi omologhi. Il numero dei cromosomi nelle cellule diploidi è caratteristico di ogni specie (nell’uomo è 46).
Interfase e divisione cellulare
Il ciclo cellulare viene distinto in due fasi: l’interfase e la divisione cellulare.
L‘interfase occupa la maggior parte del ciclo cellulare e viene divisa in 3 momenti: nella fase G1, che segue la precedente divisione cellulare, la cellula cresce di dimensioni, moltiplica gli organuli e svolge le sue funzioni caratteristiche;
nella fase S avviene la sintesi del DNA e di conseguenza la duplicazione dei cromosomi; nella fase G2 la cellula organizza le strutture necessarie alla divisione.
La divisione cellulare si compone di due processi distinti: la divisione nucleare (o mitosi o cariocinesi) e la divisione del citoplasma (o citodieresi).

Fasi della mitosi e della meiosi
La divisione cellulare nei procarioti
Nei procarioti la divisione cellulare si svolge con modalità un poco differenti
poiché il cromosoma è unico e anulare e manca la membrana nucleare. Il cro-
mosoma, attaccato a un punto della membrana plasmatica, si duplica e le due copie di cromosomi identici si attaccano alla membrana in due punti vicini. La cellula si allunga e tra i
punti di attacco si inserisce nuova membrana plasmatica che allontana i due
cromosomi. All’altezza della regione mediana della cellula, la membrana plasmatica si approfondisce verso l’interno fino a dividere la cellula madre in due cellule figlie.
La mitosi
La mitosi è l’insieme dei processi che portano alla corretta trasmissione dei cromosomi alle due cellule figlie. Nonostante il processo sia continuo, si usa distinguere 4 fasi: profase, metafase, anafase e telofase.
Nella profase i cromosomi, già duplicati durante l’interfase, si ispessiscono e diventano ben visibili. Ogni cromosoma appare formato da due cromatidi uniti tramite il centromero. La membrana nucleare e i nucleoli si dissolvono. Si forma il fuso mitotico, costituito da fibre di microtubuli che dalle due estremità della cellula (poli cellulari) convergono verso il centro. Nelle cellule animali il fuso mitotico origina dai due centrioli.
La metafase è caratterizzata dall’allineamento dei cromosomi su un piano (piano equatoriale) al centro della cellula e dal legame che si crea tra i filamenti del fuso mitotico e i centromeri. Alla fine della metafase ogni centromero si sarà diviso, per cui ogni cromatidio ne possiede uno proprio.
Durante l’anafase i filamenti del fuso “tirano” ogni cromatidio di un cromosoma in direzioni opposte verso uno dei due poli. In corrispondenza di ogni polo si trova quindi lo stesso numero di cromatidi, che coincide con quello dei cromosomi inizialmente presenti nella cellula madre.
Alla telofase i cromatidi si despiralizzano per formare l’intreccio di filamenti tipico dell’interfase. Intanto scompare il fuso mitotico e ciascun nucleo figlio viene separato dal citoplasma per mezzo di una membrana nucleare.
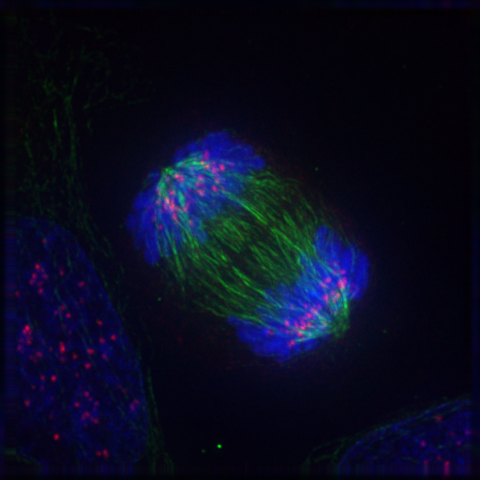
Cellula in anafase
La citodieresi
La citodieresi è il processo con cui si divide il citoplasma delle due cellule figlie. Avviene con due modalità differenti nelle cellule animali e nelle cellule vegetali. Nella zona equatoriale delle cellule animali compare una strozzatura che si fa sempre più profonda fino a determinare la separazione delle cellule figlie. Nelle piante la parete cellulare si forma a spese di una placca cellulare che compare nella regione centrale del fuso.
La riproduzione sessuale
Si definisce riproduzione sessuale la formazione di un nuovo individuo dall’unione di due cellule sessuali, provenienti da ciascuno dei due genitori.
Le cellule che prendono parte alla riproduzione sessuale sono i gameti: vengono detti spermatozoi quelli maschili, cellule uovo quelli femminili. A differenza delle cellule somatiche, i gameti sono cellule aploidi, cioé contengono un solo cromosoma. Durante la riproduzione lo spermatozoo si fonde con la cellula uovo (fecondazione) e si forma nuovamente una cellula diploide, lo zigote, che andrà incontro a ripetute divisioni mitotiche per costituire l’organismo completo. La presenza di cellule aploidi è indispensabile affinché il numero di cromosomi di una specie rimanga costante. Infatti, se lo spermatozoo e la cellula uovo fossero diploidi, di generazione in generazione il numero dei cromosomi aumenterebbe. Il meccanismo che porta alla formazione delle cellule aploidi è la meiosi.
La meiosi
La meiosi è il processo che riduce a metà il corredo cromosomico di una cellula diploide, dando origine a due cellule figlie aploidi.
La meiosi, preceduta dall’interfase durante la quale i cromosomi vengono duplicati, consiste in due divisioni cellulari, una successiva all’altra, dette meiosi I e meiosi II, che vengono divise ognuna in 4 fasi: profase I, metafase I, anafase I, telofase I e profase II, metafase II, anafase II e telofase II.
Durante la meiosi I i cromosomi, composti ciascuno da due cromatidi, formano coppie di cromosomi omologhi, che si appaiano. I cromatidi dei cromosomi omologhi si intrecciano in certi punti (chiasmi) e si verifica il crossing-over, cioè lo scambio di segmenti corrispondenti di DNA. Al termine i cromosomi omologhi di ogni coppia si separano l’uno dall’altro (segregazione), e ciascuna cellula figlia riceve uno solo dei due cromosomi di ogni coppia. Poiché la segregazione di una coppia di omologhi è indipendente dalla segregazione di un’altra coppia, si dice che avviene un assortimento indipendente dei cromosomi.
Dopo la meiosi I le cellule figlie entrano nell’interfase, durante la quale si accrescono, ma non duplicano i cromosomi. Nella meiosi II i cromatidi di ogni omologo si separano e diventano cromosomi indipendenti. In totale la meiosi dà origine a 4 cellule aploidi, i gameti, che diventeranno spermatozoi o cellule uovo a seconda che appartengano rispettivamente a un organismo maschile o femminile.
La variabilità genetica
Gli organismi che si riproducono per via sessuale sono geneticamente unici grazie alla variabilità genetica, per cui i figli ereditano dai genitori nuove combinazioni di geni e si determinano differenze genetiche tra gli organismi.
La variabilità genetica è il prodotto di mutazioni, crossingover, segregazione, assortimento indipendente e fecondazione.
Le mutazioni sono errori nella duplicazione del DNA che provocano una variazione nei nucleotidi del DNA o un cambiamento nella disposizione dei geni sui cromosomi. In entrambi i casi ciò si traduce con la produzione di una proteina differente.
Il crossing-over può avvenire in un punto qualsiasi del cromosoma e più fenomeni di crossing-over si possono verificare contemporaneamente in diversi punti dello stesso cromosoma. Le variazioni possibili sono moltissime e si originano cromosomi diversi da quelli dei genitori.
Per la segregazione e l’assortimento indipendente, il numero di combinazioni possibili di cromosomi nei gameti è dato da 2n, dove nè il numero di coppie di cromosomi. L’uomo ha 23 coppie di cromosomi omologhi, perciò può produrre 2 23 = 8.388.608 gameti diversi.
La fecondazione, cioè la fusione dei gameti di due individui diversi, offre un numero di zigoti praticamente infinito (8.388.608 x 8.388.608 = oltre 70.369 miliardi).