L’eredità delle caratteristiche biologiche
L’eredità dei caratteri
Fin dai tempi antichi, allevando animali e coltivando piante l’uomo ha avuto modo di osservare le somiglianze e le differenze esistenti tra genitori e figli. Ha così scoperto la possibilità di migliorare le razze e i raccolti, selezionando e accoppiando tra loro le varietà di animali e piante che presentavano le caratteristiche più utili. Tuttavia, pur avendo scoperto che i caratteri morfologici e fisiologici si trasmettono da una generazione all’altra, il meccanismo con cui si compie tale trasmissione è rimasto sconosciuto fino alla fine del sec. XIX e solo recentemente è divenuto oggetto di studio di una scienza specifica, la genetica.
Che cos’è la genetica
La genetica è la scienza che studia i meccanismi dell’eredità attraverso i quali avviene la trasmissione delle caratteristiche biologiche (caratteri) da una generazione a quella successiva nelle varie specie animali e vegetali.
In particolare, la genetica studia il genoma, cioè il corredo di informazioni genetiche dei vari organismi, i cui componenti sono i cromosomi e i geni, le unità funzionali ereditarie contenute in questi ultimi. I geni sono segmenti di DNA dai quali dipende la sintesi di una determinata proteina e quindi la comparsa di un dato carattere ereditario e vengono trasmessi da una generazione a quella successiva attraverso la riproduzione.
La genetica classica nasce nella seconda metà dell’800 con gli studi compiuti dal biologo boemo Gregor Mendel su piante di pisello e si sviluppa nei primi decenni del ‘900 grazie a esperimenti condotti sul moscerino dell’aceto.
Negli anni ‘40, in seguito al riconoscimento che il DNA costituisce la base fisica dell’eredità, prende avvio la genetica molecolare. A partire dagli anni ‘70, con l’impiego delle tecniche della biologia molecolare, si è aperta la strada alla manipolazione del materiale genetico e quindi allo sviluppo dell’ingegneria genetica.
La rilevanza della genetica nei diversi settori della biologia è testimoniata dallo sviluppo di discipline specialistiche, quali la citogenetica (la genetica a livello cellulare), la genetica dei microrganismi e delle piante, la genetica dello sviluppo e la genetica umana.
| CARATTERE DOMINATE | CARATTERE RECESSIVO | |
| forma del seme | liscio | rugoso |
| colore del seme | giallo | verde |
| forma del bacello | liscio | rugoso |
| colore del bacello | verde | giallo |
| colore del fiore | porpora | bianco |
| posizione del fiore | all'ascella delle foglie | all'apice dei rami |
| altezza della pianta | alta | bassa |
I sette caratteri studiati da Mendel
Gli incroci di Mendel
La scelta di Mendel delle piante di pisello per i suoi esperimenti non fu casuale. Egli aveva infatti osservato che queste piante differiscono tra loro per vari caratteri, ciascuno
dei quali compare in due forme diverse. Inoltre per la struttura stessa del fiore le piante di pisello si autoimpollinano (il polline feconda l’ovulo del suo stesso fiore): le piante che si formano mantengono le caratteristiche del fiore originario, non essendoci apporto di materiale ereditario da parte di altre piante. Quando l’autoimpollinazione si verifica per numerose generazioni, si forma una linea pura.
Mendel incrociò piante di pisello di due linee pure (P) che differivano tra loro per un solo carattere (incrocio monoibrido). I prodotti di ogni incrocio, cioè la prima generazione filiale (F1), furono piante tutte uguali, che presentavano il carattere di uno solo dei due genitori. Mendel chiamò ibridi i discendenti ottenuti dall’incrocio di due linee pure.
Incrociando tra loro le piante della generazione F1, Mendel ottenne la seconda generazione filiale (F2), costituita per tre quarti da piante che continuavano a presentare il carattere di F1 e per un quarto da piante nelle quali ricompariva il carattere presente nella generazione P, apparentemente scomparso nella F1. Mendel chiamò questo carattere recessivo (dal latino recedere, restare indietro), mentre chiamò dominante il carattere che compariva nei tre quarti delle piante.
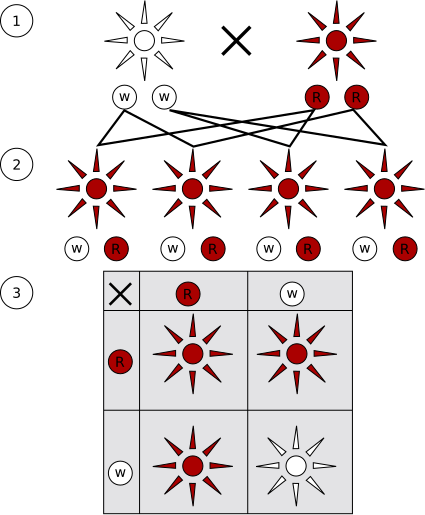
Fenotipi dominanti e recessivi. (1) Generazione parentale. (2) Generazione F1. (3) Generazione F2.
Mendel eseguì anche incroci diibridi (cioè considerò due caratteri contemporaneamente), ottenendo dei risultati analoghi: nella F1 si manifestava solo la variante dominante di entrambi i caratteri; nella F2 ottenne piante per 1⁄16 con le due varianti recessive, per 9⁄16 con le due varianti dominanti, per 3⁄16 con una variante dominante, per 3⁄16 con l’altra variante dominante.

Incroci diibridi: I fenotipi dei due tratti indipendenti nella seconda generazione mostrano un rapporto 9:3:3:1; in questo esempio il colore del pelo è indicato da B (marrone, dominante) e b (bianco, recessivo), mentre la lunghezza della coda è indicata da S (corta, dominate) e s (lunga); quando i genitori sono omozigoti per ogni tratto (SSbb e ssBB), la loro prole nella generazione F1 è eterozigote in tutti i loci e mostrano solo fenotipo dominate (SsbB); Se i figli si incrociano fra loro, nella generazione F2 appariranno tutte le combinazioni di colore del pelo e lunghezza della coda: 9 sono marroni/corte (riquadri viola), 3 sono bianchi/corte (riquadri rosa), 3 sono marroni/lunghe (riquadri blu) e uno è bianco/lunga (riquadro verde)
Mendel, il fondatore della genetica

Il biologo, matematico e monaco agostiniano ceco Gregor Mendel
Gregor Mendel (1822-1884) era un monaco del monastero di Brno (nell’attuale Repubblica Ceca), con una vasta cultura matematica acquisita all’università di Vienna. Per dieci anni, dal 1854 al 1864, egli coltivò e studiò i meccanismi dell’eredità di circa 28.000 piante di Pisum sativum, dalle quali era stato attratto per la presenza di alcuni caratteri che si presentavano in due forme alternative (il colore del seme, giallo o verde; il bacello, liscio o rugoso; i fiori, ascellari o apicali ecc.).
Mendel non fu il primo a compiere studi di questo tipo, ma ebbe alcune intuizioni che gli permisero di introdurre degli elementi di novità: la scelta di un numero elevatissimo di individui per seguire gli esperimenti nel corso di numerose generazioni; l’analisi di un solo carattere per volta; l’interpretazione statistica dei risultati; l’uso di simboli per esprimere in modo chiaro e schematico gli incroci.
L’opera di Mendel, sebbene pubblicata e inviata alle principali società scientifiche
del tempo, fu ignorata fino al 1900, quando tre studiosi di scienze biologiche, l’olandese H. de Vries, il tedesco K.E. Correns e l’austriaco E. Tschermak, l’uno indipendentemente dall’altro, riscoprirono i risultati di Mendel e ne riconobbero l’importanza.
Le tre leggi di Mendel
Al termine degli esperimenti, Mendel arrivò alle seguenti conclusioni (che gli permisero di enunciare le tre leggi di Mendel che sono alla base della genetica):
- i caratteri non si mescolano negli ibridi ma mantengono la propria identità
- ogni carattere è controllato da una coppia di “fattori” ereditari, che vengono trasmessi, uno da ciascun genitore, ai figli attraverso i gameti. Oggi si sa che questi fattori sono i geni, che sono presenti sul cromosoma in una delle due forme alternative, dette alleli, delle quali una (allele dominante) prevale sull’altra (allele recessivo), mascherandone la presenza nella F1
- al momento della meiosi, ciascuna coppia di cromosomi (uno di origine materna e uno paterno) si separa in modo che in un gamete vada solo un cromosoma; ogni spermatozoo e ogni cellula uovo possiede quindi un solo allele per ogni carattere
- con la fecondazione i gameti si combinano a caso e si riformano le coppie di cromosomi (e quindi di alleli);
- si definiscono omozigoti gli individui che hanno i due alleli di un carattere uguali (dominanti o recessivi), eterozigoti gli individui che hanno i due alleli diversi (uno dominante e uno recessivo): gli omozigoti possono produrre un solo tipo di gamete, gli eterozigoti due;
Le leggi di Mendel valgono sia per le piante sia per gli animali: anche nell’uomo molti caratteri sono trasmessi secondo queste leggi. Il bruno dei capelli è dominante sul rosso; i capelli crespi dominano su quelli lisci; gli occhi scuri su quelli azzurri; il naso aquilino su ogni altro tipo di naso.
Legge della dominanza dei caratteri o della uniformità degli ibridi
Incrociando tra loro individui che differiscono per un solo carattere, si ottengono alla prima generazione ibridi tutti uguali. Indicando gli alleli con le lettere dell’alfabeto e precisamente con A il carattere dominante e con a il carattere recessivo, nell’incrocio di due linee pure si avrà:
| generazione parentale P | AA x aa |
| gameti di P | A A a a |
| prima generazione filiale F1 | tutti Aa |
Legge della segregazione
Alla seconda generazione, ottenuta incrociando tra loro gli ibridi della prima, gli alleli che controllano un determinato carattere si separano (segregano) e vengono trasmessi a gameti diversi. Si ottengono 1⁄4 degli individui con il carattere recessivo e 3⁄4 con il carattere dominante. Di questi ultimi 2⁄3 sono eterozigoti, 1⁄3 è omozigote.
| prima generazione filiale F1 | Aa x Aa |
| gameti di F1 | A a A a |
| seconda generazione filiale F2 | AA Aa aA aa |
Si definisce fenotipo il complesso dei caratteri visibili di un individuo; genotipo la combinazione di alleli posseduta da un individuo. Da questi incroci si osserva che il fenotipo dominante è espresso sia dagli omozigoti dominanti sia dagli eterozigoti. Per determinare il genotipo dell’individuo con fenotipo dominante si ricorre al test-cross (reincrocio o incrocio di controllo) che utilizza l’omozigote recessivo.
Se il genitore con fenotipo dominante è eterozigote, i discendenti avranno per metà il fenotipo dominante e per metà quello recessivo. Se invece il genitore con fenotipo dominante è omozigote, i discendenti avranno tutti il fenotipo dominante.

Se il genitore con fenotipo dominante è eterozigote (Yy) i suoi discendenti avranno per metà il fenotipo dominante (Yy) e per metà quello recessivo (yy); se invece è omozigote dominante (YY), avranno tutti fenotipo dominante (Yy)
Legge dell’assortimento indipendente
Incrociando individui che differiscono tra loro per due o più caratteri, ogni coppia di alleli per ciascun carattere viene ereditata in maniera del tutto indipendente dall’altra. Si hanno così tutte le possibili combinazioni degli alleli di ciascuna coppia e la comparsa di individui con caratteri nuovi.
Incrociando tra loro 2 diibridi RrGg, ogni individuo dà origine a 4 tipi di gameti (RG, Rg, rG e rg) che possono combinarsi in 16 modi diversi.
Le eccezioni alle leggi di Mendel
Alcuni caratteri non mostrano la classica relazione dominanza-recessività, poiché nessuno dei due alleli domina sull’altro. In questo caso entrambi gli alleli sono espressi negli individui della F1, che non assomigliano a nessuno dei due genitori ma presentano un carattere intermedio.
Un esempio di dominanza incompleta si trova nei fiori della bocca di leone, in cui il colore dei petali è controllato da due alleli: uno per il colore rosso ®, l’altro per il colore bianco (R’). (Poiché non esiste l’allele recessivo, negli incroci di dominanza incompleta si usa una diversa simbologia.)
Dall’incrocio RR x R’R’ non si ottengono ibridi con fiori rossi o bianchi, bensì con fiori rosa RR’. Gli alleli per il colore dei fiori della F 1 non si sono modificati, tanto che nella F2 ricompaiono fiori rossi e fiori bianchi.
La legge dell’assortimento indipendente vale solo per i geni che si trovano su cromosomi diversi. Infatti i geni che si trovano sullo stesso cromosoma (geni concatenati) tendono a essere ereditati insieme: quanto più due geni sono vicini sul cromosoma, tanto maggiori sono le probabilità che siano trasmessi insieme.
Esistono alcuni caratteri che sono determinati da più di due alleli. Per esempio, il tipo di gruppo sanguigno di un individuo è determinato da 3 alleli: IA, IB, I0. Gli alleli IA e IB sono codominanti, poiché entrambi si manifestano nel fenotipo dell’eterozigote, e sono entrambi dominanti su I0. Dalle combinazioni dei 3 alleli si ottengono i seguenti fenotipi:
| GENOTIPO | FENOTIPO |
| IAIA | gruppo A |
| IAI0 | gruppo A |
| IBIB | gruppo B |
| IBI0 | gruppo B |
| IAI0 | gruppo AB |
| I0I0 | gruppo 0 |
La determinazione del sesso
Nell’essere umano, come negli altri vertebrati e in molti insetti, esiste una coppia di cromosomi dall’aspetto caratteristico. Nella femmina questa coppia è costituita da due cromosomi uguali a forma di bastoncino (cromosomi X), mentre nel maschio i due cromosomi sono diversi: uno è a forma di bastoncino, come nella femmina, l’altro è più piccolo e con un’estremità a uncino (cromosoma Y). Questa coppia è definita con il nome di cromosomi sessuali.
Al momento della formazione delle uova e degli spermatozoi, i cromosomi della coppia si separano e ciascun gamete ne riceve uno solo. Le femmine producono uova tutte uguali, contenenti sempre un cromosoma X, mentre i maschi producono due diversi tipi di spermatozoi: metà contenenti il cromosoma X e metà il cromosoma Y. Al momento della fecondazione, quando l’uovo si unisce a uno spermatozoo contenente il cromosoma X, si riforma la coppia XX e il nuovo organismo sarà femmina; se invece l’uovo si fonde con uno spermatozoo portatore del cromosoma Y, si forma una coppia XY e il discendente sarà maschio.
Il sesso di un bambino viene dunque stabilito fin dal momento della fecondazione e dipende dal tipo di spermatozoo che feconderà l’uovo. È quindi il padre a determinare il sesso del nascituro e non la madre, come si credeva in passato.
Le basi molecolari dell’eredità
Mendel aveva fatto luce sui meccanismi che regolano la trasmissione dei caratteri ereditari. Il passo successivo era analizzare la struttura fine del gene. Con una serie di brillanti osservazioni ed esperimenti, gli scienziati hanno stabilito che i geni sono localizzati sui cromosomi e che ogni gene è una porzione della molecola di DNA.
Oggi i genetisti esplorano la struttura dei geni fin nei minimi dettagli per raggiungere essenzialmente due obiettivi: chiarire il modo in cui i geni esercitano il loro controllo sulle caratteristiche di un organismo e come i geni modificano la loro attività in funzione delle necessità della cellula. Attualmente il genoma di un organismo può essere modificato grazie alle tecniche della biotecnologia.
Sebbene sia ancora molto distante, uno dei principali obiettivi della biotecnologia è correggere i difetti del genoma umano, sostituendo i geni difettosi oppure affiancandone altri normali.
La teoria cromosomica dell’eredità
La teoria cromosomica dell’eredità, secondo la quale i cromosomi sono la sede dei geni, si fonda sui seguenti presupposti:
- la stretta analogia tra il comportamento dei cromosomi durante la meiosi e quello dei geni, previsto dalle leggi di Mendel
- la presenza dei cromosomi nei gameti
- la presenza di un cromosoma materno e di uno paterno nelle cellule somatiche.
Diverse sono le prove a sostegno di questa teoria emerse da studi sul moscerino dell’aceto. Il biologo statunitense T. Morgan (1866-1945), sperimentando numerosi incroci dell’insetto, dimostrò l’esistenza di un legame fisico tra il gene responsabile del colore dell’occhio del moscerino e un determinato cromosoma.
Un altro tipo di indagine fu compiuta sui cromosomi giganti delle ghiandole salivari dell’insetto, che per le loro dimensioni e per il fatto di essere sempre despiralizzati sono ben visibili al microscopio ottico, dove appaiono costituiti da bande alternate chiare e scure. Dalla loro osservazione si poté appurare che l’assenza ereditaria di certi geni è associata alla scomparsa fisica di determinate bande.
La natura dei geni
Una volta stabilito che i geni sono localizzati sui cromosomi, il passo successivo era quello di stabilire da quale dei due costituenti dei cromosomi (DNA e proteine) è composto il gene. Per molto tempo si pensò che le proteine fossero il materiale ereditario, ma in seguito l’attenzione si indirizzò sul DNA. Gli studi che portarono all’identificazione del DNA come costituente chimico dei geni iniziarono negli anni ‘20 con gli esperimenti di Griffith e trovarono conferma con le dimostrazioni di Avery e di Hersey e Chase.
Gli esperimenti di Griffith
Nel 1928 il batteriologo F. Griffith condusse una serie di esperimenti sul batterio Streptococcus pneumoniae, l’agente della polmonite nei topi, che esiste in due forme: una virulenta, protetta da una capsula liscia e viscosa; l’altra innocua, priva di capsula e con la superficie ruvida.
Griffith osservò che poteva rendere innocui gli Streptococcus virulenti uccidendoli con il calore. Tuttavia, iniettando nei topi un miscuglio di Streptococcus virulenti uccisi con il calore e Streptococcus innocui, i topi contraevano la polmonite: nel loro sangue venivano trovati batteri virulenti vivi.
Griffith concluse che esiste un “principio trasformante” capace di indurre la trasformazione dei batteri innocui in batteri virulenti. Inoltre questa trasformazione poteva essere trasmessa ereditariamente.
Gli esperimenti di Avery
Nel 1944 lo scienziato americano O. Avery, insieme ai suoi collaboratori, dimostrò che il “principio trasformante” individuato da Griffith è il DNA.
Egli trattò con enzimi proteolitici (enzimi che rompono i legami peptidici) un miscuglio di Streptococcus virulenti uccisi con il calore e Streptococcus innocui vivi e osservò che avveniva la trasformazione. Invece, trattando il miscuglio con enzimi che distruggono il DNA, la trasformazione non aveva luogo.
Gli esperimenti di Hersey e Chase
Nel 1952 gli scienziati americani A. Hershey e M. Chase realizzarono degli esperimenti su un ceppo di virus, i batteriofagi o fagi T2 , che infetta i batteri. Essi volevano stabilire quale parte del virus reca con sé le istruzioni necessarie per la produzione di nuove particelle virali all’interno del batterio.
Ai ricercatori era noto che:
- quando un batterio viene infettato da un fago T2 , la cellula batterica si rompe e si liberano numerosi fagi;
- i fagi sono costituiti unicamente da un rivestimento proteico e da DNA;
- la molecola di DNA contiene fosforo; le proteine contengono zolfo.
Hershey e Chase aggiunsero fosforo radioattivo al mezzo di coltura di una popolazione di fagi, e zolfo radioattivo a quello di un’altra popolazione. Si formarono così un ceppo con DNA marcato (cioè contenente l’elemento radioattivo) e un ceppo con proteine marcate. Quando i batteri venivano infettati dal ceppo di virus con proteine marcate, lo zolfo radioattivo veniva trovato solo nel rivestimento dei fagi al di fuori dei batteri. Non risultavano radioattivi né i batteri, contenenti il materiale genetico dei fagi, né i nuovi fagi. Quando invece i batteri venivano infettati dal ceppo con DNA marcato, il fosforo radioattivo veniva trovato all’interno sia dei batteri infetti, sia dei nuovi fagi prodotti dall’infezione. Ciò dimostrava che il materiale genetico iniettato dai fagi nei batteri è DNA e non una proteina.
I geni influenzano le caratteristiche di un organismo
L’insieme dei geni contenuti in un singolo corredo di cromosomi di un organismo si dice genoma. Ogni gene influenza il fenotipo, cioè il complesso dei caratteri visibili di un individuo, controllando una tappa specifica di una via metabolica.
Nella maggior parte dei casi i geni controllano la sintesi di proteine (ipotesi un gene-una proteina); altri geni hanno una funzione regolatrice nei confronti di geni che codificano le proteine.
L’ipotesi un gene-una proteina
La stretta relazione tra gene e proteina è stata dimostrata negli anni ‘40 dagli scienziati americani G. Beadle ed E. Tatum, con una serie di esperimenti su un fungo ascomicete (Neurospora crassa), in grado di crescere su un terreno di coltura minimo (contenente solo sali minerali, saccarosio e la vitamina biotina). Partendo da questi precursori, il fungo sintetizza gli altri composti organici di cui necessita. Beadle e Tatum irradiarono alcuni campioni di Neurospora con raggi X, che produssero una mutazione nel loro patrimonio genetico. Gli scienziati osservarono che i funghi mutanti non erano più in grado di crescere sul terreno di coltura minimo, poiché avevano perso la capacità di far avvenire una reazione metabolica catalizzata da un determinato enzima: ne conclusero che la sintesi dell’enzima in questione doveva essere sotto il controllo del gene mutato. I risultati di queste osservazioni si possono riassumere nella teoria un gene-un enzima. Dal momento che un enzima è chimicamente una proteina, è lecito formulare la teoria nella forma un gene-una proteina.
Le mutazioni
Una mutazione è un cambiamento nella successione o nel numero delle basi di un gene. Una mutazione può avvenire in qualsiasi cellula di un organismo, ma solo quella che si verifica nei gameti entra a far parte del patrimonio ereditario dell’individuo e si trasmette da una generazione all’altra. Le mutazioni possono interessare singoli nucleotidi (mutazioni geniche) o tratti di cromosoma (mutazioni cromosomiche).
Le mutazioni geniche, dette anche puntiformi, possono insorgere spontaneamente o per rari errori durante la replicazione del DNA. Possono anche essere prodotte da alcuni composti chimici e da radiazioni. Esse comprendono:
- la delezione (perdita di un nucleotide);
- l’inserzione (aggiunta di un nucleotide);
- la sostituzione (una base sostituisce un’altra base).
Le mutazioni cromosomiche sono causate da rotture all’interno dei cromosomi, spontanee o provocate in seguito a esposizione a raggi X o ad agenti chimici. Le estremità rotte tendono a ricongiungersi provocando dei cambiamenti nell’ordine dei geni. Comprendono:
- la traslocazione (scambio di segmenti di cromosomi tra cromosomi non omologhi);
- l‘inversione (rotazione di 180° di un segmento di cromosoma);
- la delezione (perdita di un segmento di cromosoma).
Le mutazioni cromosomiche possono anche alterare il numero dei cromosomi (mutazioni genomiche): si ha polisomia o monosomia a seconda che uno o più cromosomi siano stati rispettivamente acquistati o persi.
Le mutazioni sono la principale fonte della variabilità genetica.
Nel corso di milioni di anni di evoluzione, la selezione naturale ha utilizzato le mutazioni per favorire lo sviluppo di organismi ben integrati nel loro ambiente: ogni nuova mutazione altera questo equilibrio e quindi ha molte più probabilità di essere dannosa, anzi-
ché utile. In questo modo ciascuna specie è andata via via perfezionando un proprio “programma” genetico altamente specializzato.
Tuttavia un cambiamento ambientale può alterare la stabilità di una popolazione. Allora una mutazione può rivelarsi vantaggiosa (perché migliora l’adattamento della popolazione al nuovo ambiente), tanto da entrare a far parte del genoma di una specie.
La regolazione dei geni
Una cellula utilizza solo una parte ridotta dei propri geni, più precisamente solo quelli che codificano la sintesi delle proteine di cui ha bisogno in quel momento.
Negli eucarioti pluricellulari, inoltre, i diversi tipi di cellule funzionano in modo differente, perché trascrivono solamente i geni che codificano le proteine necessarie alla loro funzione.
La cellula modula l’espressione dei propri geni attraverso meccanismi di regolazione, che si esplicano specialmente attraverso l’azione di molecole, come proteine regolatrici, o molecole segnale come gli ormoni. La spiegazione del meccanismo di controllo dei geni nelle cellule procarioti si basa sul fatto che il DNA è spesso organizzato in “strutture”, dette operoni, in cui i geni che svolgono funzioni affini (geni strutturali) sono situati l’uno vicino all’altro.
Un operone è formato da quattro parti:
- un gene regolatore che codifica una proteina, detto repressore;
- un promotore, una regione di DNA che viene riconosciuta dalla RNA-polimerasi e segna il punto di inizio della trascrizione
- un operatore, che dirige l’accesso della RNA-polimerasi al promotore per la trascrizione e al quale può legarsi il repressore (bloccando in tal modo la trascrizione)
- i geni strutturali
Nei procarioti (batteri) il funzionamento dei geni è sotto il controllo di alcune sostanze presenti nell’ambiente circostante e che fungono da effettori.
Gli effettori positivi provocano l’induzione della trascrizione di alcuni geni, o insieme di geni, mentre quelli negativi ne provocano la repressione. Di conseguenza si verifica un aumen-
to o una diminuzione della sintesi dei prodotti di tali geni.
Il compito dell’effettore è legarsi al repressore e modificarne la conformazione, così da alterarne la capacità di legarsi con l’operatore.
Se l’effettore è positivo, questa capacità diminuisce, il repressore non si lega con l’operatore, e la RNA-polimerasi non si blocca nella trascrizione del DNA. Se invece l’effettore è negativo, il repressore si lega all’operatore e la trascrizione da parte della RNA-polimerasi è bloccata.
Anche negli eucarioti vi possono essere meccanismi di regolazione a breve termine come quello dei procarioti; ma intervengono anche meccanismi più complessi a lungo termine, che si esplicano a diversi livelli. I controlli conformazionali riguardano la distinzione tra le zone di cromatina attiva nella trascrizione (eucromatina, meno condensata) e quelle che non vengono trascritte (eterocromatina, zone più compatte); il controllo trascrizionale è effettuato da fattori di trascrizione o proteine regolatrici che attivano la trascrizione di un gene; il controllo post-trascrizionale consiste nella rimozione degli introni; il controllo della traduzione e della stabilità del DNA regola il numero delle copie di mRNA e la loro durata; il controllo post-traduzionale modifica l’attivazione delle catene polipeptidiche e la durata delle proteine.

Trascrizione dell'operone
La ricombinazione genetica
La costituzione genetica di un organismo può essere modificata in modo casuale e “naturale”, oppure artificialmente in modo mirato. Il primo fenomeno avviene tra individui della stessa specie per mezzo del crossing-over.
Nei batteri, inoltre, può avere luogo tra cellule diverse grazie ai processi di trasduzione, coniugazione e trasformazione. Se invece la struttura genetica di un organismo viene alterata artificialmente, si parla di biotecnologia.
La ricombinazione genetica nei batteri
La trasduzione è operata da un fago che trasferisce una parte del DNA di un batterio a un’altra cellula batterica:
un fago che ha infettato un batterio può incorporare porzioni del suo DNA e introdurlo nel cromosoma di un altro batterio che ha successivamente infettato.
Nella coniugazione il DNA passa da un batterio a un altro con cui viene a contatto mediante un filamento di citoplasma. Le cellule prodotte dalla divisione del batterio che ha ricevuto il DNA avranno un cromosoma riassortito, poiché deriva anche dall’altro batterio.
La trasformazione consiste nel passaggio da un batterio a un altro di un plasmide, cioè del DNA supplementare rispetto al proprio cromosoma; il plasmide porta i geni che consentono di sviluppare la resistenza agli antibiotici.
Le biotecnologie
Le biotecnologie sono quelle tecniche che permettono di produrre sia sostanze sia servizi (per esempio, analisi, depurazione delle acque ecc.) con l’impiego di organismi viventi, cellule e loro costituenti, o mediante la loro manipolazione genetica.
Propriamente, le tecniche che consentono di alterare il patrimonio genetico degli organismi vanno sotto il nome di ingegneria genetica. A differenza della ricombinazione naturale, l‘ingegneria genetica altera in modo mirato il genoma di un organismo, isolando alcuni suoi geni (eventualmente modificati) e inserendoli in cellule di un’altra specie, dove si moltiplicano e sintetizzano le loro proteine. La cellula ospite (nella quale introdurre i geni) è normalmente un batterio. I vantaggi nell’uso dei batteri sono molteplici: semplicità della cellula; conoscenza dei meccanismi di ricombinazione naturale; velocità di crescita.
Una delle metodologie più importanti dell’ingegneria genetica ricorre al DNA ricombinante. Questa tecnica utilizza gli enzimi di restrizione, capaci di tagliare il DNA di due organismi (per es. batterico e umano) in corrispondenza di precise sequenze, in modo tale che le estremità delle due catene siano complementari e possano appaiarsi. Altri enzimi (DNA-ligasi) uniscono tra loro le estremità dei frammenti.
Mediante vettori di clonazione, i più comuni dei quali sono i plasmidi, il DNA ricombinante è trasferito in una cellula batterica, dove produce la proteina corrispondente al gene umano inserito. A mano a mano che il batterio si riproduce per divisione cellulare, il DNA viene amplificato, cioè riprodotto in un gran numero di copie, ottenendo clone di cellule geneticamente identiche, dotate del DNA ricombinante.
Dato che le cellule batteriche non sono in grado di rimuovere gli introni, presenti nella maggior parte dei geni degli eucarioti, e quindi non possono esprimere il gene inserito, si utilizza di preferenza il cDNA (DNA complementare); questo è ottenuto mediante la trascrittasi inversa, un enzima estratto dai virus a RNA che catalizza la trascrizione
“inversa”, da un mRNA maturo a DNA. Si forma una molecola ibrida mRNA-cDNA; poi altri enzimi rimuovono l’RNA e ricostruiscono il DNA.
Un metodo alternativo alla clonazione per ottenere grandi quantità di DNA ricombinante utilizza la reazione a catena della polimerasi (PCR).
La polimerasi è un enzima in grado di sintetizzare sequenze di macromolecole partendo da monomeri. Per esempio, la DNA-polimerasi sintetizza DNA a partire dai nucleotidi. La tecnica della reazione a catena della polimerasi riesce ad attivare in provetta lo stesso processo di replicazione del DNA che avviene nella cellula.
Le biotecnologie sono utilizzate da millenni per produrre vino, formaggio e in generale i prodotti fermentati. Hanno però assunto grande importanza solo negli anni ‘80 dopo che, con la scoperta del DNA e del codice genetico, si è compreso il funzionamento dei geni e si è perfezionata la tecnica del DNA ricombinante. Attualmente le biotecnologie trovano impiego nella cura della salute (produzione di sostanze che sono alla base di farmaci), nell’alimentazione (lavorazione dei prodotti, controllo della qualità e dello stato di conservazione degli alimenti), nella difesa dell’ambiente (trattamento dei rifiuti, depurazione delle acque e nell’agricoltura (inserimento di particolari geni utili nel genoma delle piante).
Sono detti organismi geneticamente modificati (OGM) od organismi transgenici tutti gli organismi (batteri, piante, animali, in teoria uomo compreso) il cui genoma contiene geni estranei, introdotti per migliorare alcune funzioni o per curare alcune malattie.
Il Progetto Genoma Umano
Il Progetto Genoma Umano è un programma internazionale di ricerca che si prefigge l’obiettivo di sequenziare i cromosomi umani, cioè identificare la sequenza dei nucleotidi cromosoma per cromosoma. Fu iniziato nel 1987 e ha coinvolto centri di ricerca di Stati Uniti, Inghilterra, Francia, Germania, Giappone e Cina, ciascuno con il compito di decifrare porzioni diverse di genoma.
La prima tappa del Progetto fu di localizzare i geni presenti su ciascuno dei cromosomi umani, per ottenerne una mappa; la seconda fase fu di sequenziare i geni, ossia trovare la successione delle basi su ciascun cromosoma.
Nel 2003 fu completato il sequenziamento dell’intero genoma umano. In realtà è stato decodificato il 99,9% dei geni che sono patrimonio comune di tutti gli individui della specie uomo; il nuovo più difficile compito sarà di studiare il restante 0,1%, che costituisce le varianti individuali.
Il genoma umano consiste di circa 40 mila geni per complessivamente 3 miliardi di coppie di basi distribuite su 46 cromosomi.
La conoscenza del genoma umano è un altro progresso della genetica, che troverà applicazione soprattutto in campo medico, nella ricerca di terapie in grado di rimediare ai geni difettosi (terapia genica).
Genetica umana
Gli studi di genetica umana rivelano che la nostra specie, dal punto di vista genetico, non possiede meccanismi differenti dalle altre specie animali con riproduzione sessuale. Perciò le conoscenze acquisite su altri animali possono essere applicate anche all’uomo, per studiare le caratteristiche individuali non attribuibili a influenze culturali e ambientali.
Per verificare la comparsa di determinati caratteri nei membri di una famiglia, i genetisti utilizzano mezzi diversi dagli incroci impiegati in studi su animali piccoli e con un breve ciclo vitale, principalmente l’analisi degli alberi genealogici. Gli obiettivi che la genetica umana si pone sono: chiarire il contributo dei geni allo sviluppo fisico individuale e dia-gnosticare e curare le malattie genetiche (terapia genica).
In questo ambito di ricerca si colloca il Progetto Genoma Umano.
Le malattie genetiche
Le malattie ereditarie possono essere dovute ad alterazioni dei geni o ad alterazioni dei cromosomi, di solito più frequenti con l’aumentare dell’età materna.
La maggior parte delle alterazioni dei geni è causata da alleli recessivi, che si manifestano solo in individui omozigoti per il gene difettoso. Per esempio, la galattosemia, cioè l’intolleranza al lattosio (lo zucchero del latte), si verifica quando un gene difettoso non è in grado di sintetizzare l’enzima che converte il galattosio (un componente del lattosio) in glucosio. In assenza dell’enzima, il galattosio non viene metabolizzato e si accumula nelle cellule, danneggiandole.
Altre malattie genetiche sono provocate da alleli dominanti. Forme morbose di questo tipo sono piuttosto rare, poiché le persone che ne sono affette muoiono spesso prima di aver raggiunto la maturità sessuale e quindi non si riproducono. Una forma che si manifesta tardivamente è invece la corea di Huntington, malattia gravissima che insorge in età adulta. È caratterizzata da movimenti tipici, detti appunto coreici (involontari, improvvisi, a scatti, presenti a riposo e nell’esecuzione di atti volontari e assenti nel sonno) e progressiva riduzione delle capacità mentali. Dato che i sintomi compaiono in età matura, la persona che ne è affetta può aver avuto figli, il 50 per cento dei quali può manifestare la malattia, prima di conoscere le sue condizioni.
Fondamentale è la prevenzione a livello di consulenza genetica (si sconsiglia di avere figli se in famiglia è presente una patologia di questo tipo).
Le anomalie cromosomiche possono interessare sia i cromosomi sessuali, o eterocromosomi, sia gli autosomi (cromosomi non sessuali).
Esempi di anomalie dei cromosomi sessuali sono la sindrome di Turner e quella di Klinefelter. La maggior parte delle persone affette da sindrome di Turner, che colpisce solo le femmine, ha 45 cromosomi invece dei normali 46: quello mancante è un cromosoma X (configurazione cromosomica X0). Le principali caratteristiche sono bassa statura, assenza o forte ritardo dello sviluppo dei caratteri sessuali secondari, assenza di mestruazioni e un certo grado di ritardo mentale. La sindrome di Klinefelter è caratterizzata dall’eccedenza di uno o più cromosomi X nelle cellule di un individuo di sesso maschile, per cui è presente una configurazione cromosomica XXY o, più raramente, XXXY. Si manifesta con comparsa di caratteri sessuali secondari femminili, sterilità e intelligenza al di sotto della media.
Infine, un numero anomalo di autosomi si forma durante la meiosi per un processo di non-disgiunzione: durante la seconda divisione meiotica la segregazione si svolge in modo anomalo e un gamete può ricevere entrambe le copie di un cromosoma, oppure nessuna. Con la fecondazione lo zigote può avere 3 copie di uno stesso cromosoma (trisomia) o una copia sola (monosomia). Circa un quarto delle gravidanze con feti portatori di anomalie cromosomiche sono interrotte da un aborto spontaneo. Il caso più noto di anomalia degli autosomi, che non influisce sul normale decorso della gestazione, è la trisomia 21, nota anche come sindrome di Down, la cui incidenza aumenta con l’aumentare dell’età materna e in caso di parente prossimo affetto da tale sindrome. Dovuta alla presenza di 3 copie del cromosoma 21, si manifesta con un aspetto fisico caratteristico (taglio degli occhi orientaleggiante, lineamenti minuti, parte posteriore della testa appiattita, mani piccole e tozze) e ritardo mentale di grado variabile.