L'Evoluzione e diversità dei viventi
L’evoluzione degli organismi
L’idea di evoluzione degli organismi è molto antica, ma, dopo secoli di abbandono, fu elaborata in modo scientifico solo nell’800, grazie all’opera di Darwin e Wallace. Basata sui concetti di variabilità, selezione naturale e lotta per l’esistenza, la teoria evolutiva fu perfezionata con lo sviluppo di nuove discipline biologiche, soprattutto della genetica: oggi è una delle teorie fondamentali della biologia.
Il concetto di evoluzione
Alla luce della teoria di Darwin e Wallace, per evoluzione si intende il processo attraverso cui le diverse specie di viventi derivano da forme di vita preesistenti, in seguito a graduali modificazioni, o variazioni geniche, trasmesse ereditariamente nel corso del tempo in base a un meccanismo determinato dalla selezione naturale: sopravvivono e si assicurano una discendenza quegli individui che presentano variazioni favorevoli, tali cioè da renderli meglio adatti a fronteggiare le condizioni ambientali.
Il pensiero evoluzionistico
Un’idea di evoluzione dei viventi era presente già nelle opere di filosofi greci e di poeti latini. Per quasi duemila anni, tuttavia, dominò incontrastato il pensiero del filosofo e naturalista greco Aristotele (384-322 a.C.), che ordinava tutti gli organismi lungo una “scala della natura”: ai gradini più bassi si collocava la materia inanimata, mentre l’ultimo gradino era occupato dall’uomo, secondo uno schema definito già dalla creazione.
Le idee di Aristotele confluirono in seguito nel pensiero cristiano, e fino all’illuminismo il pensiero scientifico rimase cristallizzato nella teoria della immutabilità delle specie (fissismo), create all’inizio da una mente divina (creazionismo).
Solo nell’800, con lo sviluppo di scienze come la geologia e la paleontologia, si riaccese la discussione sull’evoluzione dei viventi. Molte erano, infatti, le prove che andavano accumulandosi in favore dell’evoluzione:
- l’esplorazione di nuove terre mostrava una grande varietà di forme viventi;
- i fossili di specie estinte mostravano molte somiglianze con organismi viventi;
- l’età della Terra, stimata fino allora in poche migliaia di anni, si rivelò molto più antica, tale da consentire un intervallo di tempo sufficiente per il realizzarsi di graduali modificazioni nelle specie.
Una delle prime teorie evoluzionistiche fu proposta dal biologo e naturalista francese J.-B. de Lamarck (1774-1829). La teoria di Lamarck si basa su due assunti principali:
- uso e non uso degli organi: gli organi di un animale non ancora adulto possono svilupparsi o atrofizzarsi in rapporto al loro minore o maggiore uso
- ereditarietà dei caratteri acquisiti: le modificazioni dei caratteri acquisiti durante la vita dell’individuo possono essere trasmesse ai figli
L’esempio più famoso addotto da Lamarck per spiegare la sua teoria è quello della giraffa. Essa avrebbe sviluppato, nel corso di generazioni, un collo lungo nel tentativo di raggiungere le foglie alte degli alberi (meno soggette a esaurirsi di quelle dei rami più bassi).
La teoria di Lamarck, oggi respinta perché non suffragata da valide prove sperimentali, ha tuttavia il merito di aver sottolineato l’importanza della variabilità dei caratteri ereditari nei meccanismi evolutivi.
La teoria dell’evoluzione di Darwin-Wallace
Una spiegazione delle cause dell’evoluzione e la formulazione di una teoria su solide basi scientifiche fu merito dei naturalisti inglesi Charles Darwin (1809-1882) e Alfred Russel Wallace (1823-1913). Contemporaneamente e indipendentemente l’uno dall’altro, essi elaborarono idee analoghe, che furono esposte nel 1858 in due saggi passati quasi inosservati.
Nel 1859 Darwin pubblicò le sue conclusioni nell’opera L’origine delle specie, che, contrariamente ai saggi precedenti, ebbe subito grande risonanza (per questo motivo e anche per la superiore statura scientifica di Darwin, la teoria dell’evoluzione, o darwinismo, è specialmente legata al suo nome).
Darwin e Wallace basavano il loro pensiero su numerose osservazioni dirette (compiute soprattutto durante viaggi in regioni tropicali), che evidenziavano in particolare:
- la grande varietà di specie presenti in una stessa regione
- il perfetto adattamento di queste specie al diverso tipo di habitat e di alimentazione.
Importanti furono inoltre gli spunti forniti da dottrine in precedenza eleaborate in ambito geologico (Lyell) e demografico-economico (Malthus). Secondo il principio dell’attualismo del geologo inglese C. Lyell (1797-1875) il modellamento della Terra è il risultato non di immani catastrofi, come terremoti o eruzioni gigantesche, ma di forze naturali lente e continue, sempre all’opera. Analogamente, in campo biologico piccole variazioni di forma da una generazione all’altra avrebbero potuto formare, nel corso del tempo geologico, tutte le specie animali e vegetali che conosciamo. Secondo la teoria demografica dell’economista inglese T. Malthus (1766-1834), le popolazioni umane tendono a crescere in progressione geometrica, mentre le risorse alimentari in natura crescono in progressione aritmetica, comportando una scarsità di risorse a danno degli individui più deboli, che soccombono. Da qui l’dea di una continua lotta per l’esistenza, generalizzabile a tutti gli organismi viventi, e il cui risultato sarebbe quello di favorire i più adatti (selezione naturale).
L’evoluzione per selezione naturale
La spiegazione dell’evoluzione di Darwin-Wallace, nota come teoria dell’evoluzione per selezione naturale, può essere così riassunta:
- fra gli individui di una stessa specie vi è grande variabilità genetica (che si manifesta in piccole differenze nei caratteri, quali corporatura, altezza, pigmentazione della pelle, colore degli occhi ecc.)
- le variazioni individuali devono essere ereditabili, perché i figli sono simili ai genitori
- tutti gli organismi tendono a moltiplicarsi, ma l’ambiente non permette una crescita indiscriminata, per cui le dimensioni di una popolazione sono frenate dalla mortalità (selezione naturale)
- sopravvivono e si riproducono più facilmente gli individui che hanno raggiunto un migliore adattamento all’ambiente in cui vivono, e che quindi sono favoriti nella lotta per l’esistenza
- con questi meccanismi, le specie nel tempo si evolvono, dando origine a nuove specie.
Darwin conosceva le tecniche della selezione artificiale, il mezzo attuato da secoli da allevatori e coltivatori per migliorare le razze economicamente utili, e ipotizzò che un meccanismo simile potesse verosimilmente agire anche in natura. Non conosceva invece le leggi dell’ereditarietà (gli studi di Mendel, suo contemporaneo, passarono quasi inosservati fino ai primi del ‘900) e non seppe quindi spiegare in particolare come si origina la variabilità di caratteri (sia fisici, sia comportamentali) sulla quale avrebbe dovuto agire la selezione naturale.
La teoria dell’evoluzione ha comunque il merito di aver sottolineato che i nuovi caratteri si originano indipendentemente dall’ambiente (cioè non è l’ambiente a creare nuovi caratteri, come sosteneva Lamarck), ma, una volta comparsi, sono selezionati dall’ambiente. L’evoluzione è quindi diretta dalla selezione naturale, ma procede in modo casuale.
La teoria dell’evoluzione ebbe grande impatto sul pensiero dell’800 e, in particolare, sulla biologia, di cui rimane ancora oggi una delle teorie unificatrici, perché permette di spiegare e di organizzare in modo logico tutte le conoscenze delle diverse discipline.

Le varie specie di fringuello delle Isole Galapagos derivano dallo stesso antenato comune
Le prove dell’evoluzione
Dopo la morte di Darwin la teoria evolutiva fu sostenuta con fermezza o fortemente contestata. Tuttavia, molti punti oscuri della teoria sono stati chiariti, e sono state confermate numerose prove a sostegno dell’evoluzione. Diverse scienze hanno contribuito a fornire le prove alla teoria dell’evoluzione: la paleontologia, l’anatomia comparata, l’embriologia, la biochimica, la biogeografia.
- La testimonianza dei fossili: il loro studio rivela una graduale successione di forme che variano nel tempo, dalle più semplici alle più complesse: gli strati rocciosi più superficiali e quindi più recenti contengono organismi più simili a quelli attuali; quelli più profondi, e più antichi, forme con maggiori differenze.

Calcare con resti fossili di bivalvi presenti come impronte del guscio
- L’analogia delle strutture: molte specie non imparentate che vivono nello stesso ambiente mostrano strutture simili nella morfologia esterna, ma di diversa origine anatomica; per esempio, le pinne delle foche (mammiferi) e dei pinguini (uccelli). Questa convergenza evolutiva dimostra che la selezione naturale ha favorito in organismi diversi, presenti in uno stesso ambiente, adattamenti simili.

Squalo (Chondrichthyes) |

Delfino (Mammalia) |

Ittiosauro (Reptilia) |
Animali marini: gli squali, appartenenti alla classe dei pesci cartilaginei, i delfini della classe dei mammiferi e gli Ittiosauri della classe dei rettili (che vissero nel mesozoico), sotto la pressione dell’ambiente acquatico che li circonda si sono evoluti assumendo una morfologia idrodinamica complessivamente molto simile.”
- L’omologia fra strutture diverse: in molte specie adattate ad ambienti diversi si osservano strutture morfologiche diverse, ma con una struttura di base simile; questo confermerebbe la derivazione da un comune antenato (per esempio, l’arto dei tetrapodi è formato dalle stesse ossa, modificate dall’adattamento nel corso dell’evoluzione in ali o pinne).

L'omologia degli arti nei vertebrati
- Presenza di strutture vestigiali, o residuali: in alcune specie si riconoscono organi molto ridotti, non più funzionali, ben sviluppati invece in altre specie. Anche in questo caso sarebbero resti di una comune struttura ancestrale non più utile all’adattamento (per esempio, nell’uomo le ossa del coccige sarebbero “avanzi” della coda)

Plica semilunare, residuo della membrana nittitante che in altri mammiferi, negli uccelli, nei rettili e negli anfibi è molto più sviluppata e può essere distesa sull'occhio, a palpebra aperta, svolgendo una funzione di protezione e lubrificazione.
- La somiglianza degli stadi embrionali precoci di animali di gruppi diversi si spiega ammettendo un lontano antenato comune

Somiglianza fra embrioni (fila in alto), fra pesci, salamandra, tartaruga, gallina, maiale, vacca, coniglio e umano
- Le basi biochimiche della vita (DNA, proteine ecc.) sono comuni a tutti gli esseri viventi: questo confermerebbe una derivazione comune di tutti i viventi da uno stesso organismo primigenio.

Un cladogramma che collega tutti i gruppi importanti di organismi viventi al LUA (il tronco (segmento) nero nella parte inferiore). Questo grafico è derivato dalla sequenza del RNA ribosomico.
- L’enorme diversità di specie esistenti: i diversi gradi di somiglianza permettono di stabilire legami evolutivi più o meno stretti e ricostruire la storia evolutiva degli organismi. Su questi caratteri comuni si basa anche la classificazione degli esseri viventi, che rispecchia lo sviluppo delle grandi linee evolutive. Oggi, le tecniche di sequenziamento del DNA permettono un confronto diretto e molto preciso dei geni di specie diverse.
- La distribuzione geografica dei viventi: alcune specie si trovano solo in una regione isolata o in un dato continente. Questo può essere spiegato dalla differente storia evolutiva delle singole specie, avvenuta dopo che sono state separate geograficamente da eventi geologici.

La biodiversità
- L’evoluzione in atto: a conferma della continuità del processo evolutivo basterebbero alcuni esempi di modificazioni di specie osservati in brevi intervalli di tempo. Per esempio, la rivoluzione industriale ha favorito le specie che si sono adattate a un nuovo ambiente, più ricco di fumi inquinanti (melanismo industriale); l’uso dei pesticidi ha favorito gli insetti resistenti; l’uso degli antibiotici la resistenza di alcuni ceppi batterici.

Evoluzione del cavallo
La teoria moderna dell’evoluzione
Grazie agli sviluppi della genetica all’inizio del ‘900 si scoprì che le caratteristiche ereditarie sono controllate dai geni, localizzati nei cromosomi della cellula.
Grande importanza fu data anche allo studio statistico della variabilità genetica di una popolazione. Il matematico inglese G.H. Hardy e il medico tedesco W. Weinberg definirono, indipendentemente, le condizioni di stabilità o di equilibrio di una popolazione: si tratta della legge di Hardy-Weinberg secondo cui, in mancanza di forze perturbatrici, cioé in condizioni di equilibrio, le frequenze relative di ciascun allele di una popolazione si trasmettono inalterate di generazione in generazione.
In natura, tuttavia, è difficile che una popolazione sia perfettamente in equilibrio, perché si verifica sempre almeno una delle seguenti condizioni “perturbatrici”:
- mutazioni
- flusso genico tra popolazioni
- dimensioni piccole
- accoppiaenti non casuali
- selezione naturale
Pertanto la legge di Hardy-Wienberg può essere una conferma in termini matematici del potenziale evolutivo di una popolazione: perché vi sia un’evoluzione, occorre infatti che nel corso delle generazioni le frequenze alleliche di una popolazione non rimangano costanti, cioè che la popolazione non sia in equilibrio.
Verso la fine degli anni ‘40, in seguito alle conoscenze della genetica, della statistica, della biologia e della genetica molecolari fu così possibile definire una formulazione sintetica della teoria dell’evoluzione: fra gli organismi di una popolazione esiste una certa variabilità individuale, provocata dall’azione costante di mutazioni e dalla ricombinazione dei geni che si verifica a ogni generazione; tale variabilità è casuale e non orientata e su di essa agisce come fattore direttivo principale la selezione naturale.
I fattori dell’evoluzione
L’evoluzione è, in sintesi, il cambiamento genetico di una popolazione. I principali fattori coinvolti nell’evoluzione sono dunque: popolazione, variabilità e selezione.
La popolazione
La popolazione è l’insieme di individui di una stessa specie che vivono in una medesima area.
Ogni individuo possiede un corredo di geni che costituisce il suo genotipo e, interagendo con l’ambiente, ne definisce il fenotipo, cioè i caratteri morfologici e comportamentali.
L’insieme dei geni (e degli alleli) di tutti gli individui di una popolazione costituisce il pool genico della popolazione; la frequenza allelica è invece la proporzione con cui i diversi alleli sono presenti nella popolazione. Compito della genetica delle popolazioni è studiare la frequenza, la distribuzione e l’ereditarietà dei geni di una popolazione.
La variabilità
In un individuo le fonti della variabilità genica sono le mutazioni e la ricombinazione genetica durante la riproduzione sessuale.
Le mutazioni sono rari cambiamenti dell’informazione genetica: molti non hanno effetti immediati sull’organismo, altri sono dannosi e pochi sono benefici: non sono quindi la causa primaria dell’evoluzione, ma sono fonte di nuovi alleli, su cui possono intervenire altri agenti dell’evoluzione.
La ricombinazione genetica si produce durante la riproduzione sessuale all’atto della meiosi, quando i cromosomi omologhi sono distribuiti a caso nei gameti; oppure durante il crossing-over, con scambio di porzioni di cromosomi; o infine all’atto della fecondazione, per l’unione casuale di un gamete maschile e uno femminile. In una popolazione, fonti di variabilità sono la migrazione e la deriva genetica.
La migrazione è intesa come flusso di geni tra popolazioni (e non solo “spostamento” fisico di una popolazione): per esempio, la “migrazione” di un maschio dominante in un nuovo branco fornisce nuovi geni al pool genico del branco. La migrazione può distribuire alleli vantaggiosi nelle diverse popolazioni di una specie e allo stesso tempo contribuisce a mantenere le caratteristiche della specie in tutto il territorio.
La deriva genetica è la fluttuazione della frequenza allelica in una piccola popolazione dovuta a un evento casuale.
Nel meccanismo detto a “collo di bottiglia” l’evento casuale è determinato da una notevole riduzione del numero degli individui della popolazione (per esempio, per malattia o carestia): se la popolazione non si estingue, la frequenza allelica delle future generazioni sarà determinata dal patrimonio dei pochi individui rimasti.
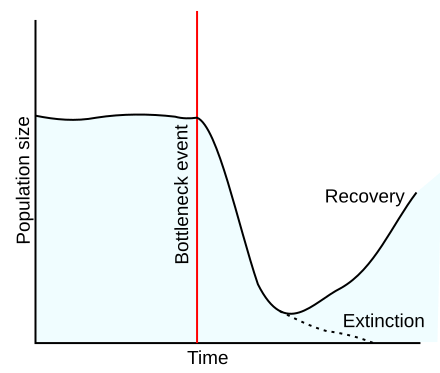
Effetto collo di bottiglia: la popolazione o riesce a sopravvivere o si estingue
Analogo è “l’effetto del fondatore“: se pochi individui colonizzano una nuova zona e rimangono isolati, da quei pochi pionieri deriverà una nuova popolazione con frequenze alleliche diverse da quella di origine. In una popolazione ampia è meno probabile che questo alteri le frequenze geniche in modo da determinare un cambiamento apprezzabile nella popolazione, cioè un’evoluzione. In una popolazione di pochi individui, invece, la frequenza di alcuni alleli è bassa, per cui avvenimenti casuali possono facilmente eliminarli dal pool genico.

Effetto del fondatore: a sinistra la popolazione originale, a destra tre possibili colonie
La selezione naturale
Uno dei meccanismi cardinali della teoria evolutiva è la selezione naturale: l’insieme dei processi che all’interno di una popolazione consentono ad alcuni individui di sopravvivere e riprodursi.
Gli individui in cui si manifesta un carattere favorevole all’ambiente possiedono un maggiore valore adattativo, o fitness, e sono dunque “selezionati”: hanno cioè più possibilità di riprodursi rispetto al resto della popolazione e nelle generazioni successive sarà presente un maggior numero di individui che possiedono quel carattere “favorevole” (riproduzione differenziale).
In questo modo la selezione naturale porta a una graduale modificazione della composizione genetica della popolazione, nella quale i caratteri con maggiore fitness si manifesteranno con frequenza sempre maggiore. Nella popolazione si verifica allora una pressione di selezione che modifica la distribuzione dei fenotipi, cambiando la posizione nella popolazione del carattere medio più frequente.
La selezione naturale non provoca cambiamenti genetici negli individui, ma, agendo sulla loro possibilità di sopravvivenza e sulla capacità riproduttiva, controlla indirettamente la variabilità genetica, per cui l’evoluzione si manifesta nella popolazione.
I cambiamenti evolutivi non sono necessariamente “buoni” o finalizzati, ma solo preferibili in quel momento e in quell’ambiente: il risultato della selezione naturale è quindi “solo” l’adattamento all’ambiente, sia alla sua componente abiotica (caratteristiche fisico-climatiche), sia a quella biotica (costituita dagli organismi). Nel riquadro a pagina 108 sono descritti alcuni meccanismi particolari di selezione.

Pavo cristatus. Un esempio di selezione sessuale: il bellissimo piumaggio esibito dal pavone maschio serve ad attrarre e orientare la scelta della femmina.
Il concetto di specie e la speciazione
La modificazione di una popolazione per selezione naturale può procedere fino al punto in cui le diversità della popolazione attuale rispetto a quella originaria saranno tali da poter considerare la popolazione attuale come una nuova specie. Dal punto di vista biologico, si definisce specie l’insieme di organismi con caratteristiche simili in grado di accoppiarsi e dare prole feconda; gli individui appartenenti a una stessa specie hanno un patrimonio genetico (pool genico) comune. Fra gli individui di una stessa specie si osserva tuttavia, come abbiamo visto, una certa variabilità: ciascuna specie possiede dunque le potenzialità per una continua evoluzione, in relazione alle modificazioni ambientali.
Alcuni meccanismi particolari di selezione
Alcuni interessanti meccanismi selettivi sono quelli che agiscono a livello delle interazioni tra organismi.
La competizione tra individui della stessa specie per le medesime risorse è uno dei meccanismi più comuni e diretti di selezione.
La coevoluzione è la continua reciproca interazione tra due specie, ciascuna delle quali esercita una pressione selettiva sull’altra. Per esempio, l’evoluzione della corolla dei fiori è stata determinata, e a sua volta ha determinato l’evoluzione degli apparati boccali degli insetti
che da essa traevano il nettare; nella predazione si instaura una coevoluzione tra predatore e preda, per cui il predatore è costretto a sviluppare nuove strategie di attacco in seguito al perfezionamento di quelle di difesa della preda (o viceversa). La selezione sessuale è la scelta del compagno attuata da alcune specie di pesci e da molte specie di uccelli e mammiferi. È evidente che alcune caratteristiche del maschio (colori appariscenti o un certo tipo di corteggiamento) preferite dalle femmine saranno determinanti del suo successo riproduttivo.
I meccanismi della speciazione
Il processo che porta alla formazione di una nuova specie prende il nome di speciazione. Si forma una nuova specie quando i componenti di una popolazione hanno subìto cambiamenti genetici (divergenza) tali per cui non possono più accoppiarsi tra loro o comunque dare prole feconda (isolamento riproduttivo).
I meccanismi della speciazione che favoriscono la divergenza sono ricollegabili all’isolamento geografico, alla radiazione adattativa e alla poliploidia.
- Isolamento geografico. Due popolazioni di una stessa specie rimangono separate da una barriera fisica per lungo tempo. Questa barriera può essere dovuta a eventi geologici di grande portata, come la deriva dei continenti, o di portata minore, come la presenza di una catena montuosa, di un fiume o di una colata lavica. Se l’isolamento persiste per lungo tempo, ogni popolazione sviluppa mutazioni differenti e quindi una differente evoluzione e, anche se ricongiunte, non saranno più in grado di accoppiarsi tra loro.
- Radiazione adattativa. Diverse popolazioni di una specie invadono nicchie ecologiche differenti e adattandosi si evolvono in nuove specie in tempi relativamente brevi. La radiazione adattativa avviene, per esempio, nel caso di colonizzazione di nuovi ambienti o quando si perfeziona un adattamento nuovo (come il sangue caldo dei mammiferi).
- Poliploidia. La poliploidia è costituita dalla presenza in alcuni individui di copie multiple di cromosomi, invece del numero diploide comune alla maggior parte delle specie animali e vegetali. Si osserva piuttosto frequentemente nelle piante, quando una cellula uovo fecondata duplica i cromosomi, ma non si divide in due cellule figlie. Gli individui poliploidi sono ibridi in genere più vigorosi dei genitori, ma sono sterili: infatti la progenie derivata dalla fecondazione di un gamete poliploide con numero doppio di cromosomi e un gamete “normale” aploide con un numero singolo di cromosomi avrà un numero dispari di cromosomi; questi non riescono ad appaiarsi, non possono portare a compimento la meiosi e di conseguenza produrre gameti. Gli individui poliploidi possono tuttavia riprodursi in modo asessuato e dare così origine piuttosto velocemente a una nuova specie.

Il martin pescatore africano pigmeo (Ispidina picta), che vive nelle foreste a sud del Sahara |

Corythornis cristatus: le due specie sono affini ma l'enorme estensione delle foreste le ha isolate diversificandole |
Il mantenimento dell’isolamento riproduttivo
La nuova specie, una volta differenziata, è importante che mantenga l’isolamento riproduttivo con le altre specie; questo può avvenire con meccanismi prezigotici, cioè che impediscono l’accoppiamento, o postzigotici, cioè che impediscono di generare prole feconda.
Esempi di meccanismi prezigotici sono:
- l‘isolamento geografico, quando vi è una barriera fisica
- l’isolamento ecologico, quando vi è un preciso adattamento a nicchie diverse
- l’isolamento comportamentale: due popolazioni della stessa specie che vivono nello stesso luogo possono sviluppare un diverso comportamento riproduttivo (diverso periodo riproduttivo, diversi richiami o rituali di corteggiamento)
- l’incompatibilità meccanica, quando due popolazioni sviluppano differenze morfologiche delle strutture riproduttive (per esempio, nelle piante la posizione di stami e pistilli nei fiori).
I meccanismi di isolamento postzigotici comprendono:
- l’incompatibilità genetica, quando lo spermatozoo non può raggiungere la cellula uovo
- la debolezza degli ibridi, che non sopravvivono
- la sterilità degli ibridi, che non si riproducono.
Le estinzioni
Gli stessi meccanismi dell’evoluzione che possono portare alla formazione di nuove specie possono condurre all’estinzione di altre, che non hanno saputo adattarsi a nuove condizioni ambientali o alla convivenza con nuove specie (per esempio, di predatori). Dall’inizio della vita sulla Terra, molte specie non sono state in grado di continuare la loro evoluzione e nel tempo sono lentamente scomparse.
In alcuni periodi geologici, tuttavia, si sono registrate estinzioni quasi contemporanee di un gran numero di specie differenti. Queste spettacolari estinzioni di massa fanno pensare ad avvenimenti catastrofici particolari: per esempio, le estinzioni della fine del Mesozoico in cui scomparvero anche i grandi dinosauri sarebbero state provocate dagli effetti della caduta di un grande meteorite.
I più comuni fattori responsabili delle estinzioni, tuttavia, sono in genere meno vistosi: innanzitutto la distribuzione localizzata e l’eccessiva specializzazione della specie, che non offrono molte possibilità di adattamento in caso di variazioni ambientali; cause più dirette di estinzione sono la competizione tra specie, la comparsa di nuovi predatori (tra cui ha avuto e ha un ruolo fondamentale l’uomo) o parassiti, la distruzione dell’habitat.
Microevoluzione e macroevoluzione
Quasi tutti i processi biologici possono essere studiati a diversi livelli. Anche per l’evoluzione si parla di una microevoluzione e di una macroevoluzione.
La microevoluzione si occupa dei cambiamenti che avvengono in tempi brevi nell’ambito di una specie (mutazioni, deriva genetica, selezione naturale). Un esempio molto studiato è la modificazione della farfalla Biston betularia in seguito all’inquinamento.
La macroevoluzione si occupa dei grandi cambiamentiche sono avvenuti in tempi lunghi: per esempio, quelli che hanno portato alla diversificazione di gruppi di specie, come la differenziazione dei vertebrati dagli invertebrati o dei mammiferi dai rettili.
A un differente livello di organizzazione, oggi si studia anche l’evoluzione molecolare, che cerca le affinità nelle diverse specie delle proteine e del DNA, le molecole più complesse e importanti negli esseri viventi.
Tutti questi studi permettono di valutare il grado di somiglianza anche a livello genetico dei diversi organismi e tracciare con sempre maggiore precisione la storia della vita sulla Terra.

Esempio di microevoluzione, Biston betularia: A. comune. B. carbonaria
La storia della vita sulla Terra
La comparsa delle prime forme viventi fu preceduta, secondo le ipotesi ritenute più attendibili, da una fase detta evoluzione prebiotica, in cui si formarono le molecole organiche necessarie alla costruzione del materiale cellulare. Dalle forme di vita ancestrali consistenti di organismi procarioti, si svilupparono nel mare organismi dotati di cellule eucarioti vegetali (alghe), capaci di fotosintesi, e animali (invertebrati marini) progenitrici delle piante e degli animali, che grazie alla presenza di ossigeno nell’atmosfera poterono a un certo punto passare sulla terraferma.
Nel percorso evolutivo degli animali, la tappa più recente è la comparsa dei mammiferi, nel cui ambito si diversificarono i primati, l’ordine al quale appartiene l’uomo. L’evoluzione dell’uomo moderno si compì attraverso un insieme di eventi detto ominazione.
Dall’evoluzione prebiotica alle prime cellule
Per evoluzione prebiotica, precedente la comparsa della vita, si intende l’evoluzione chimica che ha portato alla formazione di composti organici partendo da molecole inorganiche. Questo processo si è svolto nel corso del primo miliardo di anni a partire dalla formazione della Terra (avvenuta circa 4600 milioni di anni fa).
Per spiegare la serie di eventi con cui si sarebbe svolta l’evoluzione prebiotica, gli scienziati hanno proposto diverse ipotesi. Secondo una ricostruzione plausibile l’atmosfera della Terra primordiale era composta in prevalenza da idrogeno, metano, ammoniaca, vapor acqueo e diossido di carbonio (anidride carbonica); l’ossigeno era invece presente in quantità trascurabile (per questo si dice che l’atmosfera era riducente). La radiazione solare e i fulmini fornivano l’energia necessaria per spezzare i legami di sole molecole inorganiche e formare molecole organiche semplici. Queste ultime, in assenza di ossigeno (con cui avrebbero reagito decomponendosi), ebbero modo di accumularsi in bacini d’acqua calda. Si formò così il cosiddetto brodo primordiale.
Successivamente, le molecole nel brodo primordiale (tra cui amminoacidi, lipidi, zuccheri semplici) si sarebbero aggregate in strutture sferiche, chiamate microsfere, ipotizzate come le prime cellule. Ogni microsfera era dotata di una membrana che separava l’ambiente interno da quello esterno; poteva assorbire del materiale (nutrimento) dall’ambiente esterno, crescere di dimensioni e replicarsi.
Schema biogeochimico dell’ecosistema dei vulcani sottomarini
L’evoluzione prebiotica riprodotta in laboratorio
Negli anni ‘20 il biochimico russo A. Oparin (1894-1980) e il biologo inglese J. Haldane (1892-1964) formularono l’ipotesi secondo cui in un’atmosfera riducente comuni reazioni chimiche possono produrre molecole organiche a partire da molecole inorganiche.
Trent’anni più tardi (1953) lo scienziato americano S. Miller (n.1930) verificò sperimentalmente quest’ipotesi. Egli ideò un’apparecchiatura nella quale i gas dell’atmosfera primitiva erano sottoposti, in assenza di ossigeno, a scariche elettriche.
Miller constatò la formazione di molecole organiche, tra cui alcuni amminoacidi.
Ancora negli anni ‘50 lo scienziato americano S. Fox riuscì a ottenere le microsfere. Egli sperimentò che, riscaldando degli amminoacidi in assenza di ossigeno, si formano spontaneamente dei polipeptidi, che egli chiamò protenoidi. Messi in acqua, i protenoidi costituiscono una specie di doppia pellicola che circonda una piccola vescicola, la microsfera.
Secondo ipotesi successive, un ruolo fondamentale sarebbe stato svolto da minerali argillosi che, essendo dotati di attività catalitica, avrebbero facilitato l’aggregazione delle semplici molecole organiche nei corrispondenti polimeri.
Altri scienziati ancora propongono la teoria della panspermia, secondo la quale la vita è approdata sulla Terra proveniente dallo spazio.

Esperimento di Miller sul brodo primordiale
Dalla cellula procariote alla cellula eucariote
In base ai resti fossili rinvenuti, le prime cellule viventi comparvero circa 3900 milioni di anni fa. Si tratta di cellule procarioti anaerobiche ed eterotrofe, riconducibili a batteri. Questi si nutrivano dei composti organici presenti nell’ambiente e ricavavano l’energia necessaria al
proprio mantenimento grazie a processi di fermentazione.
Il progressivo impoverimento delle sostanze nutritive nel brodo primordiale favorì quelle cellule che, per sintetizzare i composti organici, avevano sviluppato la capacità di attuare la fotosintesi, cioè utilizzare il diossido di carbonio quale fonte alternativa di carbonio e la radiazione solare per ricavare l’energia necessaria a questo processo. Comparvero così insieme alla fotosintesi le prime cellule autotrofe, i cianobatteri.
La trasformazione fotosintetica dell’acqua e del diossido di carbonio in composti organici libera ossigeno come sottoprodotto. L’atmosfera iniziò così ad arricchirsi di ossigeno e si crearono le condizioni favorevoli per l’evoluzione dei batteri aerobici, cioè in grado di effettuare la respirazione cellulare, un processo più efficiente della fermentazione per ricavare energia dagli alimenti.
Secondo le ipotesi circa 1,6 miliardi di anni fa alcuni batteri anaerobici, che si nutrivano inglobando batteri interi, avrebbero dato origine alle cellule eucarioti. Ciò sarebbe avvenuto in seguito al verificarsi di due tipi di simbiosi metaboliche:
- tra un batterio anaerobico ed eterotrofo e un batterio aerobico (che si evolve in mitocondrio): in questo caso si sarebbe formata la cellula animale;
- tra un batterio contenente dei mitocondri e un cianobatterio (che si evolve in cloroplasto): in questo caso si sarebbe formata la cellula vegetale.
È probabile che anche i flagelli e altri organuli cellulari siano stati acquisiti con questo tipo di simbiosi. La membrana nucleare, invece, si sarebbe formata in seguito a un ripiegamento all’interno della membrana cellulare a formare una cavità.
| ERA | PERIODO | EPOCA | ANNI FA (milioni) | EVENTI BIOLOGICI PRINCIPALI |
| Precambriano | 4600-3500
3500-590 |
Nascita delle prime cellulae viventi; dominio dei batteri; comparsa della fotosintesi e sviluppo di ossigeno nell'atmosfera; prime alghe e invertebrati marini a corpo molle | ||
| Paleozoico | Cambriano | 590-505 | Diffusione delle alghe marine primitive; sviluppo della maggior parte dei tipi di invertebrati marini | |
| Ordoviciano | 505-438 | Predominio degli invertebrati marini, in particolare artropodi e molluschi; primi pesci, funghi; invasione delle terre emerse a opera delle piante | ||
| Siluriano | 438-408 | Molti pesci, trilobiti e molluschi nelle acque oceaniche; prime piante vascolari; invasione della terraferma da parte degli artropodi | ||
| Devoniano | 408-360 | Diffusione dei pesci e trilobiti nei mari; comparsa di anfibi e insetti | ||
| Carbonifero | 360-286 | Foreste palustri di felci arboree e licopodi; dominio degli anfibi; numerosi insetti; comparsa dei rettili | ||
| Permiano | 286-248 | Prime gimnosperme; massice estinzioni marine, compresi gli ultimi trilobiti; grande sviluppo dei rettili e declino degli anfibi | ||
| Mesozoico | Triassico | 248-213 | Primi mammiferi, dinosauri e altri rettili, foreste di gimnosperme e felci arboree | |
| Giurassico | 213-144 | Dominio dei dinosauri e conifere; primi uccelli; angiosperme | ||
| Cretaceo | 144-65 | Dominio delle angiosperme; estinzione in massa di forme di vita marine e di qualcuna terrestre, inclusi gli ultimi dinosauri | ||
| Cenozoico | Terziario | Paleocene | 65-54 | Diffusione di uccelli, mammiferi (tra cui i primati), insetti e angiosperme; Driopitecine e Ramapitecine |
| Eocene | 54-37 | |||
| Oligocene | 37-24 | |||
| Miocene | 24-5 | |||
| Pliocene | 5-2 | |||
| Quaternario | Pleistocene | 2-0,01 | Evoluzione di Homo; estinzione di molti mammiferi di grosse dimensioni | |
| Olocene | 0,01-oggi |
Orologio geologico: una proiezione dei 4,5 miliardi di anni della Terra sotto forma di orologio ("Ma" = Megaanno = un milione di anni fa; "Ga" = Gigaanno = un miliardo di anni fa
L’evoluzione nel mare
L’evoluzione sarebbe proseguita nel mare, poiché l’atmosfera non aveva ancora le caratteristiche per consentire la sopravvivenza dei viventi.
La ricostruzione degli eventi che hanno segnato il passaggio da organismi unicellulari a organismi pluricellulari è ancora incerta. I vantaggi offerti dall’organizzazione pluricellulare sono: l’aumento delle dimensioni (una singola cellula, al contrario, deve mantenersi piccola perché il rapporto superficie-volume sia ottimale), la specializzazione, la riproduzione sessuata.
I primi organismi pluricellulari erano privi di parti dure adatte alla fossilizzazione, perciò di loro si sono conservate solo alcune impronte. Si sa che circa 1,4 miliardi di anni fa comparvero le alghe e 600 milioni di anni fa i primi animali, simili a meduse.
Circa 590 milioni di anni fa si verificò un cambiamento nella composizione dell’acqua del mare: l’arricchimento di silice e sali di calcio e fosforo favorì lo sviluppo di strutture mineralizzate (gusci e conchiglie) che meglio si conservano come fossili. Da questo momento i resti fossili sono perciò più frequenti e denotano la presenza in mare di organismi
dalla forma varia (trilobiti, ammoniti, “vermi” e altri di dubbia classificazione).
Circa 500 milioni di anni fa comparvero gli ostracodermi, i primi animali dotati di una struttura di sostegno interna (endoscheletro). Gli ostracodermi erano animali privi di mascelle e di arti, ricoperti da una corazza ossea da cui usciva solo la coda. Da questi più tardi si svilupparono i pesci.

Si ritiene che l'alga verde Volvox aureus sia molto simile alle prime piante pluricellulari
Il passaggio sulla terraferma
Con il progressivo aumento dell’ossigeno nell’atmosfera e la formazione dello strato di ozono a schermo delle radiazioni ultraviolette (dannose per le cellule perché provocano mutazioni genetiche), l’ambiente della terraferma divenne idoneo all’insediamento dei viventi.

Ricostruzione museale (Museo statale di Storia Naturale di Stoccarda), tetrapode primitivo e uno dei primi vertebrati a possedere delle zampe riconoscibili, anatomicamente intermedio tra i pesci dalle pinne lobate e i primi tetrapodi pienamente capaci di camminare sulla terra
L’evoluzione delle piante
I primi organismi che si svincolarono dall’acqua furono vegetali derivati dalle alghe verdi, che gradualmente svilupparono strutture simili a radici (per l’ancoraggio alla superficie), vasi (per il trasporto dell’acqua e della linfa) e organi adibiti alla riproduzione (per proteggere i gameti dalla disidratazione).
Il clima caldo-umido che caratterizzò un lungo periodo (360-286 milioni di anni fa) favorì lo sviluppo di felci arboree e licopodi, i cui resti fossilizzati hanno prodotto i vasti giacimenti di carbone fossile.
Un successivo periodo secco (286-248 milioni di anni fa) segnò il passaggio alle gimnosperme (sono gimnosperme, per esempio, pini e larici), resistenti alla siccità; da queste 100 milioni di anni più tardi evolsero le angiosperme (con i semi racchiusi nel frutto), che attualmente dominano l’ambiente terrestre.

Evoluzione delle piante: Charophyta, muschi, felci, gimnosperme e angiosperme
L’evoluzione degli animali
Poco dopo le piante anche gli animali iniziarono a colonizzare la terraferma.
Per primi gli artropodi (crostacei, ragni, insetti ecc.), che già nel mare avevano sviluppato una struttura di sostegno esterna (esoscheletro) impermeabile e rigida. Non più sostenuto dall’acqua, l’esoscheletro evitava al corpo di essere “schiacciato” dalla forza di gravità. Gli artropodi svilupparono forme varie e di grandi dimensioni, che dominarono incontrastate l’ambiente terrestre per decine di milioni di anni.
Circa 350 milioni di anni fa dai crossopterigi, un gruppo di pesci dotati di pinne robuste e muscolose e di un’estroflessione del canale digerente (che in caso di necessità poteva essere impiegato come rudimentale polmone), derivarono gli anfibi. Gli anfibi svilupparono veri e propri arti e polmoni più efficienti. Essi però non si svincolarono definitivamente dall’acqua, dalla quale dipendevano soprattutto per la riproduzione.
Da un gruppo di anfibi, che avevano sviluppato degli adattamenti alla siccità, derivarono i rettili. Diversamente dai loro progenitori, i rettili hanno la pelle coperta di squame impermeabili, la loro fecondazione è interna e le uova sono protette da un guscio impermeabile. Come gli anfibi, anche i rettili sono animali ectotermi, o eterotermi, cioè disperdono facilmente il calore corporeo, per cui la loro temperatura non è costante ma varia con quella dell’ambiente. I rettili dominarono sulla fauna terrestre per circa 250 milioni di anni. Durante questo periodo si svilupparono forme di grandi dimensioni, tra le quali i dinosauri. Alcuni rettili si adattarono alla vita acquatica, altri divennero capaci di compiere voli planati, grazie a particolari espansioni della pelle.
Circa 65 milioni di anni fa un mutamento climatico su scala planetaria, provocato probabilmente dall’impatto di un enorme meteorite con la Terra, determinò l’estinzione di numerose specie, tra cui tutte le ammoniti e molti rettili.
Nel frattempo alcuni rettili avevano sviluppato peli e penne per trattenere il calore corporeo (animali endotermi, od omeotermi). In questo modo essi potevano essere attivi anche di notte ed evitare i predatori ectotermi, che erano intorpiditi dal freddo.
Gli uccelli derivano da rettili provvisti di penne, che ne svilupparono di più lunghe e forti alle estremità degli arti anteriori e della coda. Poterono così dapprima planare e in seguito volare, grazie anche ad alcuni adattamenti dello scheletro e della muscolatura.
Dai rettili ricoperti di peli si svilupparono i mammiferi, caratterizzati dal fatto di partorire figli vivi e non uova (viviparità) e dalla presenza di ghiandole mammarie. I primi mammiferi erano di piccole dimensioni e attivi solo di notte.
Quando la maggior parte dei rettili si estinse, i mammiferi si diversificarono e occuparono tutti gli ambienti.

Scheletri di Allosaurus (a sinistra) e di Stegosaurus (a destra)
L’evoluzione dell’uomo
L’insieme degli eventi attraverso cui si è compiuta l’evoluzione della specie umana viene detto ominazione.
Le prime tappe dell’ominazione si svolsero in Africa. Tradizionalmente si fa iniziare l’ominazione 70 milioni di anni fa, quando dai mammiferi si svilupparono i primati, l’ordine al quale appartiene l’uomo. I primati originari erano animali notturni, arboricoli, con mani e piedi prensili, muso piatto;
gli occhi erano grandi e frontali e consentivano una visione stereoscopica (percezione della profondità). Progressivamente alcuni primati divennero diurni, acquisirono la capacità di vedere i colori e per spostarsi da un ramo all’altro adottarono la brachiazione. Questo metodo di locomozione, che consiste nel tenersi sospeso con le braccia, comportò delle modifiche a livello della colonna vertebrale e del bacino, che consentirono la successiva evoluzione della stazione eretta, tipica dell’uomo.
A circa 20 milioni di anni fa risale un gruppo di primati, le Driopitecine, con caratteri umanoidi, ma vita ancora arboricola. Dalle Driopitecine derivarono le Ramapitecine, vissute da 14 a 8 milioni di anni fa. La loro comparsa avvenne quando un lungo periodo freddo determinò il ritiro della foresta tropicale e lo sviluppo di savane, un tipo di vegetazione che male si accorda con la vita arboricola. In effetti, la dentatura delle Ramapitecine rivela un parziale adattamento alla vita al suolo.
La comparsa degli ominidi
Si definiscono ominidi i progenitori dell’uomo con stazione eretta e locomozione bipede. I primi ominidi appartengono al genere Australopithecus e i loro resti fossili sono stati trovati in Tanzania e in Etiopia. Essendo bipede, Australopithecus non aveva necessità di impiegare l’arto superiore per la locomozione; questo poté allora perfezionarsi nella manipolazione e nella presa di oggetti. Gli Australopithecus comparvero attorno a 4,4 milioni di anni fa con la specie A. ramidus, della quale conserviamo solo alcuni denti. Di un’altra specie, A. afarensis, è stato scoperto nel 1974 lo scheletro quasi completo di una femmina, che è stata battezzata dagli scopritori Lucy.
Circa 2 milioni di anni fa, da qualche popolazione di un’altra specie, A. africanus, ebbe origine Homo abilis, capace di produrre rudimentali utensili in pietra scheggiata.
Quasi contemporaneamente comparve in Africa una nuova specie, forse derivata da H. abilis: Homo ergaster, uomo piuttosto alto e slanciato che viveva nella savana. Probabilmente H. ergaster migrò dall’Africa in Asia, dove lasciò come discendenza Homo erectus (1,8 milioni di anni fa), più progredito nella fabbricazione degli utensili. H. erectus si spostò dalle originarie terre africane in Eurasia; imparò a utilizzare il fuoco per scaldarsi e forse per cuocere i cibi, a cacciare grandi animali, a stabilirsi in accampamenti. Sviluppò il linguaggio, che favorì l’organizzazione degli uomini in tribù.

Australopithecus africanus ricostruzione
L’uomo attuale
Circa 200.000 anni fa in Europa si trovava Homo neanderthalensis, derivato forse da un ramo di H. ergaster parallelo a H. erectus. Con l’uomo di Neanderthal aumentò il sentimento sociale: le tribù, meglio strutturate, avevano tradizioni proprie, curavano gli anziani e seppellivano i morti; svolgevano forse pratiche rituali.
Circa 90.000 anni fa, comparve l’uomo moderno, Homo sapiens, che un poco alla volta assimilò le popolazioni dell’uomo di Neanderthal. H. sapiens era in grado di lavorare ossa e corna da cui otteneva, tra l’altro, aghi e arpioni per la pesca. Sviluppò un senso artistico, testimoniato da numerosi ritrovamenti di pitture rupestri, statuette di animali e figure femminili. Addomesticò gli animali e sviluppò l’agricoltura.

L'uomo di Neanderthal, ultimo congenere noto del sapiens è convissuto almeno per alcune decine di migliaia di anni con la nostra specie. L'immagine mostra una ricostruzione fatta dal Landesmuseum für Vorgeschichte Sachsen-Anhalt ad Halle, Germania: la tesi principale, esposta nel 2006, confermata nel 2007 e basata su tecniche avanzate di biologia molecolare, ipotizza che la specie, in Europa, abbia sviluppato individui di carnagione bianca con capelli rossi; il tipo di pigmentazione è in accordo con la scarsa irradiazione ultravioletta del territorio colonizzato.
| A. afarensis | A. africanus | H. habilis | H. ergaster | H. erectus | H. neanderthalensis | H. sapiens | |
| volume cranico
(cm3) |
400 | 450-500 | 680-750 | 1000 | 800-1300 | 1300-1600 | 1300-1600 |
| altezza
(cm) |
100 F
### M |
120 | 150 | 160-170 | 160-170 | 150-160 | 160-180 |
| dentatura | mista tra scimmia e uomo; spazi tra denti; canini sporgenti | canini normali; denti contigui; molari grandi | denti propozionati ma più grandi di quelli di H. sapiens | denti proporzionati | denti piccoli, ma poco più grandi di quelli di H. sapiens | canini proporzionati, molari grandi | denti più piccoli dei precedenti |
| fronte | sfuggente | inclinata | inclinata | quasi dritta | quasi dritta | inclinata | dritta |
| mento | assente | assente | assente | accennato | accennato | accennato | presente |
| cibo | frutti secchi,
granaglie |
frutti secchi,
granaglie, bacche, uova e larve |
carne di animali morti | vegetali, carne | vegetali,
carne |
vegetali,
carne |
vegetali,
carne |
| abitudini | raccolta | raccolta | raccolta di vegetali e animali morti | raccolta,
caccia |
raccolta,
caccia |
raccolta,
caccia |
raccolta,
agricoltura, caccia domesticazione degli animali |
Principali caratteristiche degli ominidi
Introduzione ai viventi: principi di classificazione
L’enorme diversità di specie animali e vegetali che si sono evolute sulla Terra in milioni di anni ha reso necessario ideare un sistema logico per identificare ogni organismo e assegnarlo a un’opportuna categoria secondo un criterio di classificazione; ciò anche in relazione alla probabilità di confrontare più facilmente i diversi organismi per individuarne, tra l’altro, le diverse correlazioni evolutive.
La classificazione degli organismi
Viene chiamata sistematica, o tassonomia (dal greco táxis: ordinamento), la scienza che studia i criteri di classificazione degli organismi viventi secondo uno schema basato su una serie di categorie sistematiche, o táxa(singolare táxon), disposte a livelli gerarchici.
Le sette categorie principali sono, in ordine gerarchico decrescente: regno, phylum (per gli animali) o divisione (per le piante e i funghi), classe, ordine, famiglia, genere e specie.
Ciascuna categoria sistematica superiore comprende tutte quelle inferiori (come se fossero “scatole cinesi”): così, il regno comprende numerosi phyla, ogni phylumpiù classi, ogni classe più ordini ecc., fino ad arrivare alle singole specie.
La nomenclatura binomia
Un organismo è identificato quando viene indicato a quale specie, genere, famiglia, ordine, classe, phylum (o divisione) esso appartiene. In pratica, però, per l’identificazione di un organismo si ricorre alle due categorie inferiori: il genere e la specie. Il nome scientifico di un organismo è infatti indicato per mezzo della nomenclatura binomia ideata nel ‘700 da Linneo. Questa nomenclatura è costituita da due nomi latini: il primo designa il genere, scritto per convenzione in maiuscolo, il secondo designa la specie, scritto in minuscolo. Così, il lupo è Canis (genere) lupus (specie), il cane Canis (genere) familiaris (specie), l’uomo Homo (genere) sapiens (specie).
Il nome di un nuovo genere o di una nuova specie viene assegnato nel corso di congressi scientifici internazionali, secondo diversi criteri: può essere il nome classico, tramandato dagli antichi greci o latini; può derivare dal nome dello scopritore; oppure indicare una caratteristica dell’esemplare.
La nomenclatura binomia viene usata come riferimento internazionale per superare la confusione che si crea con la varietà dei nomi con cui ciascuna lingua, e quasi ogni dialetto, designa nell’uso comune i diversi animali e piante.
La tassonomia moderna
Nell’800, con la diffusione delle teorie evoluzionistiche e del nuovo concetto di specie (un insieme di individui che possono incrociarsi fra loro e la cui prole risulta a sua volta fertile), prevalsero i sistemi di classificazione che tengono conto delle relazioni filogenetiche (o evolutive) tra le diverse specie, cercandone le derivazioni da antenati comuni.
Oggi la tassonomia ricerca con sempre maggiore precisione le affinità evolutive degli organismi e si avvale dello studio di nuove discipline: oltre all’anatomia e all’embriologia, grande importanza ha la biochimica. Quest’ultima, tra l’altro, permette di determinare la sequenza di amminoacidi delle proteine; poiché le proteine sono codificate dal DNA, quanto maggiore è la somiglianza nella composizione proteica tra due specie differenti, tanto più è probabile che i loro rapporti evolutivi siano stretti.
La tassonomia è una scienza complessa e la sistematica dei diversi raggruppamenti, soprattutto degli organismi inferiori, è tuttora controversa. I táxa sono soggetti a continue revisioni in seguito ai risultati delle ricerche nei diversi campi.
I regni dei viventi
Attualmente quasi tutte le classificazioni concordano sulla divisione degli organismi nei seguenti cinque regni: monere (Monera), protisti (Protista), piante (Plantae), funghi (Fungi), animali (Animalia), comprendenti ciascuno organismi con alcune caratteristiche fondamentali comuni.
- le monere sono procarioti unicellulari
- i protisti sono eucarioti autotrofi ed eterotrofi unicellulari e pluricellulari con tessuti non differenziati
- le piante sono eucarioti pluricellulari autotrofi
- i funghi sono eucarioti pluricellulari eterotrofi (saprofiti)
I virus non rientrano in nessuno di questi regni e vengono sempre considerati a parte, al limite del mondo vivente.
| Regno | MONERE | PROTISTI* | PIANTE | FUNGHI | ANIMALI |
| Phylum | Archeobatteri
Eubatteri |
Euglenofite
Bacillariofite Dinoflagellati Rodofite Feofite Clorofite Sarcomastigofori Sporozoi Ciliati Mixomiceti Acrasie |
Rodofite
Feofite Clorofite Briofite Pteridofite Coniferofite Antofite |
Zigomiceti
Oomiceti Ascomiceti Basidiomiceti Deuteromiceti |
Poriferi
Cnidari Platelminti Nematodi Anellidi Artropodi Molluschi Echinodermi Cordati |
I cinque regni dei viventi ( sono indicati solo i principali phyla di ogni regno)
Gli alberi evolutivi
La storia filogenetica dei raggruppamenti sistematici superiori è per comodità schematizzata in raffigurazioni grafiche, con linee che partono dai progenitori più antichi e sintetizzano il loro percorso evolutivo fino alle specie attuali.
Come abbiamo visto, tuttavia, la storia evolutiva della Terra e dei suoi abitanti, dalla formazione della vita a oggi, è un processo complesso e per molti versi conosciuto solo per via ipotetica. Pertanto non è possibile tracciare un percorso lineare dell’evoluzione, in quanto la maggior parte delle linee evolutive è interrotta da estinzioni o si divide in nuove specie.
Più spesso, dunque, gli schemi evolutivi, e quindi gli schemi tassonomici che li rispecchiano, assomigliano a grandi alberi ramificati, i cosiddetti alberi evolutivi: il tronco indica gli antenati comuni, i grossi rami le grandi divisioni del mondo vivente e le ramificazioni più piccole gli ulteriori percorsi evolutivi dei diversi gruppi di organismi.
I cicli vitali
Il ciclo vitale è l’intera sequenza delle fasi dello sviluppo di un organismo, dalla cellula fecondata (lo zigote) alla produzione delle cellule sessuali (i gameti) e alla riproduzione.
Il ciclo vitale di un organismo è considerato un elemento importante per valutare il grado di evoluzione della specie a cui appartiene e quindi per la sua classificazione.
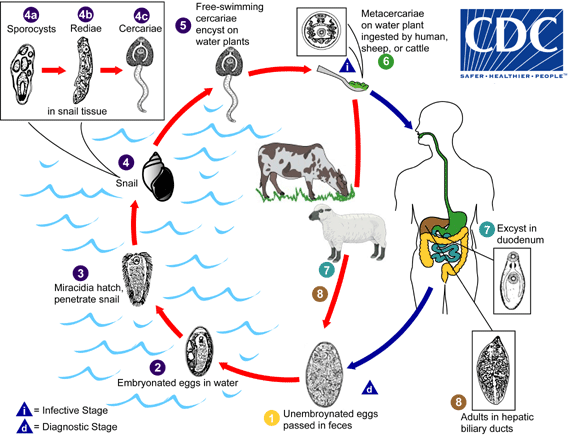
Ciclo vitale di Fasciola hepatica e suoi diversi ospiti
Le generazioni
La fase del ciclo vitale di un organismo vivente è caratterizzata da divisioni di cellule in cui viene mantenuto lo stesso numero di cromosomi, ed è detta generazione.
Gli organismi che si riproducono asessualmente per semplice divisione (mitosi) mantengono sempre lo stesso numero di cromosomi nelle cellule e il ciclo quindi si esaurisce tra due successive divisioni e conosce una sola generazione.
Negli organismi a riproduzione sessuata la produzione di gameti implica invece un dimezzamento, in seguito alla meiosi, del numero di cromosomi di alcune cellule (che si indica con n) e un raddoppiamento del numero dei cromosomi dei gameti in seguito alla fecondazione (2n). In questo caso dunque il ciclo vitale è costituito da un’alternanza di generazione aploide (cioè con metà numero di cromosomi, compresa tra la formazione delle cellule sessuali, cioè la meiosi, e la fecondazione) e di generazione diploide (con numero di cromosomi doppio, compresa tra la fecondazione e la meiosi).
Caratteristica di alcuni gruppi di organismi e indice di minore o maggiore evoluzione è il prevalere di un tipo di generazione sull’altra. Si possono così distinguere organismi aplonti (con prevalenza della generazione aploide, come molti protisti, alghe e funghi), diplonti (con prevalenza della generazione diploide, come gli animali) e aplodiplonti (con equilibrio tra le due generazioni come molte piante).
In molte piante la meiosi produce spore aploidi che si dividono per mitosi producendo un individuo aploide, o gametofito; il gametofito produce due gameti che si fondono in uno zigote diploide; questo formerà un individuo diploide, o sporofito, che a sua volta produrrà in seguito a meiosi spore aploidi per ricominciare il ciclo. Nel corso dell’evoluzione la fase gametofitica si va riducendo e nelle piante superiori predomina lo sporofito diploide (la maggior parte delle piante che vediamo), mentre il gametofito è ridotto a poche cellule del fiore per la produzione dei gameti.
Nel ciclo vitale di alcuni animali diplonti, come i celenterati, l’alternanza delle generazioni avviene tra uno stadio a larva natante (medusa), che produce i gameti e si riproduce sessualmente, e l’adulto ancorato al fondale (polipo) che si riproduce in modo asessuato.

Ciclo aplodiplonte
I virus
I virus sono particelle subcellulari, prive cioè di una struttura cellulare, capaci di moltiplicarsi soltanto all’interno di una cellula ospite da loro infettata. Per le loro ridotte dimensioni, che variano dai 17 ai 3000 nanometri, i virus possono essere osservati soltanto al microscopio elettronico.
Tutti i virus sono costituiti da un involucro proteico (capside) formato da tante subunità (capsomeri) di una stessa proteina. Alcuni virus possiedono all’esterno del capside uno strato di protezione lipidico che il virus asporta dalla membrana della cellula infettata, quando, alla fine del ciclo infettivo, la cellula si rompe e libera le nuove unità di virus replicatesi. All’interno dell’involucro vi è un acido nucleico, che può essere DNA o RNA.

Batteriofagi in azione.
Classificazione dei virus
I virus sono classificati, secondo il tipo di acido nucleico presente, in DNA virus o RNA virus. In base agli organismi che vanno a infettare, si possono dividere in virus veri e propri, che possono infettare cellule eucarioti, e in virus batteriofagi o fagi, che infettano cellule procarioti batteriche.
I virus veri e propri hanno generalmente forma di icosaedro (un poliedro a venti facce) e includono i virus del raffreddore, delle verruche, della poliomielite. I virus parassiti di cellule vegetali hanno invece per lo più forma allungata o sferica.
I fagi hanno una struttura più complessa, in cui si possono riconoscere cinque parti, costituite da altrettante proteine: testa, stilo, guaina contrattile, piastra basale, fibre della coda.
Alcuni dei virus più diffusi.
Cicli infettivi
I virus si riproducono solo all’interno di una cellula ospite: l’involucro proteico è specializzato per legarsi a un dato gruppo molecolare sulla superficie di una specifica cellula ospite.
Alcuni virus – per esempio i fagi – iniettano nella cellula ospite solo l’acido nucleico virale; in altri casi, il virus penetra interamente nella cellula e successivamente si libera dell’involucro, che può essere digerito dagli enzimi dell’ospite.
Il materiale genetico del virus si integra con quello dell’ospite, che è costretto in questo modo a “leggere” anche le informazioni genetiche virali e a produrre quindi i diversi componenti virali che verranno poi assemblati rapidamente; i nuovi virus fuoriescono dalla cellula, distruggendola, e invadono le cellule vicine.
In alcuni DNA virus, il doppio filamento di DNA iniettato nella cellula viene duplicato immediatamente e il processo si conclude con la disgregazione, o lisi, della cellula e la formazione dei nuovi virus (ciclo litico). In altri virus, il DNA integrato in quello dell’ospite rimane quiescente per un certo tempo, partecipando inattivamente alla crescita e alla divisione della cellula (ciclo lisogenico); solo successivamente avvia un ciclo litico con la distruzione della cellula.
Negli RNA virus il materiale genetico, prima di poter essere introdotto nella cellula ospite, deve essere copiato in una molecola di DNA: questo avviene mediante un enzima, detto trascrittasi inversa.
Uno dei più noti RNA virus è il retrovirus HIV, responsabile della sindrome da immunodeficienza acquisita, o AIDS.
Un tipico ciclo di replicazione virale
Le monere e i protisti
Le monere, o batteri, e i protisti sono gli organismi viventi con il livello di organizzazione cellulare più semplice: sono infatti costituiti da una sola cellula o da più cellule indifferenziate.
Questa è di tipo procariote nelle monere, di tipo eucariote nei protisti.
In entrambi i gruppi si osserva già la tendenza dell’evoluzione all’aggregazione: in alcuni casi gli organismi unicellulari si uniscono in colonie, dove iniziano a differenziare le loro funzioni.
Il regno delle monere
Le monere, o batteri, sono microscopici organismi unicellulari procarioti. Le loro dimensioni vanno fino a 2 micron di diametro e 100 micron di lunghezza: sono quindi più grandi dei virus, ma molto più piccoli delle cellule eucarioti.
Per la loro grande varietà di adattamenti, i batteri sono gli esseri viventi più numerosi sulla Terra, diffusi in tutti gli ambienti, compresi quelli caratterizzati da condizioni estreme, come l’assenza di ossigeno, le alte temperature o l’elevata salinità.
Composizione di immagini di diversi procarioti
Struttura
Come in tutte le cellule procarioti, in quelle dei batteri non vi è un nucleo distinto e delimitato da una membrana nucleare; i ribosomi sono piccoli e mancano gli organuli.
Nei batteri il materiale genetico è costituito da un’unica molecola di DNA circolare libera nel citoplasma (cromosoma batterico); vi possono anche essere piccoli anelli di DNA detti plasmidi.
La membrana plasmatica, che racchiude il citoplasma, è ricoperta di una parete cellulare rigida (ma non contenente cellulosa come nelle cellule vegetali). In alcuni batteri, la parete cellulare è avvolta da uno strato mucoso, detto capsula, costituito da polisaccaridi. Alcuni batteri hanno uno o più flagelli (privi però di microtubuli, come hanno le cellule eucarioti) o strutture filiformi relativamente rigide, dette pili o fimbrie, che servono per fissarsi a certi substrati.
I batteri possiedono diversi tipi di forme caratteristiche: le più comuni sono quella a bastoncino dei bacilli, quella sferoidale dei cocchi, quella a spirale degli spirilli e quella a virgola dei vibrioni. Inoltre, possono essere isolati o uniti in catenelle o filamenti.

Esempio tipico di cellula procariote
Batteri gram-positivi e gram-negativi
Con la tecnica di colorazione di Gram (dal nome del batteriologo danese H.C. Gram che la descrisse nel 1884) è possibile distinguere due tipi di pareti cellulari, in base alle quali si distinguono batteri gram-positivi e batteri gram-negativi. La parete cellulare dei batteri gram-negativi include una membrana che talvolta è tossica per i mammiferi e causa alcune malattie.
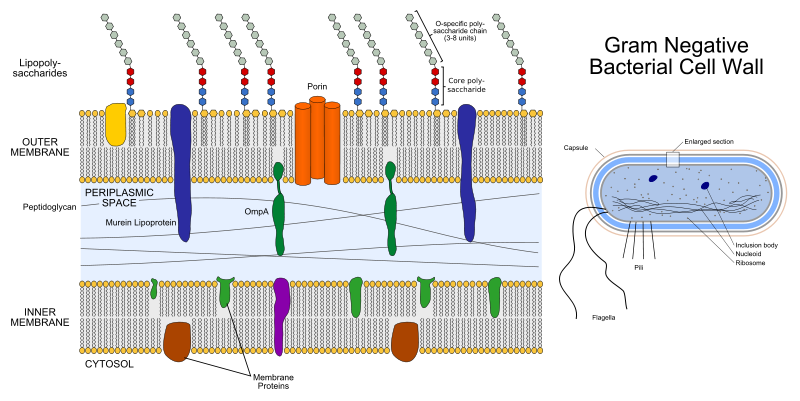
Struttura della parete cellulare di un Gram negativo
Riproduzione batterica
In condizioni ottimali i batteri si riproducono velocemente per divisione cellulare (scissione binaria) ogni 15-45 minuti.
Si osserva anche una forma di sessualità, detta coniugazione batterica, per cui due batteri si scambiano materiale genetico, in genere plasmidi, senza tuttavia dividersi.
Alcuni batteri formano spore, forme protettive o di resistenza, chiamate anche endospore (poiché sviluppate all’interno della cellula), in grado di sopravvivere in condizioni ambientali avverse ed essere trasportate per lunghe distanze nell’aria o nell’acqua. Le spore, che contengono materiale genetico e alcuni enzimi, germineranno quando si troveranno in condizioni favorevoli.
Forme batteriche
Evoluzione ed ecologia dei batteri
I batteri sono i più antichi organismi conosciuti: risalgono a circa 3,5 miliardi di anni fa. Grazie alla semplicità di organizzazione e alla mancanza di competizione con gli eucarioti, hanno avuto enorme diffusione, adattandosi in tutti gli ambienti naturali (terreno, acque), oltre che in ambienti “particolari” costituiti da animali e piante (sia all’esterno, sia all’interno di essi).
I batteri sono estremamente specializzati, perché una volta adattati al loro particolare ambiente non possono vivere in altre condizioni.
Alcuni batteri possono vivere solo in presenza di ossigeno e sono detti aerobi obbligati; altri, detti anaerobi obbligati, non possono vivere in presenza di ossigeno; altri ancora, infine, possono vivere sia in presenza, sia in assenza di ossigeno e sono detti anaerobi facoltativi.
Per quanto riguarda la temperatura, i batteri psicrofili vivono solo a temperature inferiori a 20 °C, i mesofili tra 20 e 45 °C, i termofili a temperature superiori ai 45 °C. I batteri possono condurre vita libera, o essere parassiti o simbionti.
In relazione al modo in cui ricavano l’energia necessaria per vivere, i batteri possono essere autotrofi (in grado di sintetizzare le sostanze organiche necessarie) oppure eterotrofi (che utilizzano sostanze organiche già elaborate). I batteri autotrofi includono forme capaci di fotosintesi al pari delle piante (come i cianobatteri) e forme capaci di chemiosintesi, cioè in grado di ricavare energia da reazioni in cui l’ossigeno si combina con composti inorganici (quali sulfuri e ammoniaca). Molti batteri eterotrofi sono decompositori e partecipano ai più importanti cicli biogeochimici.
Alcuni batteri sono azotofissatori: sono aerobici, in genere flagellati, capaci di utilizzare l’azoto atmosferico per la sintesi di sostanze organiche azotate. Alcuni sono a vita libera, altri (Rhizobium) vivono nei tubercoli radicali delle leguminose, realizzando una forma di simbiosi.
Altri batteri simbionti vivono nell’apparato digerente di numerosi animali, uomo compreso (nel quale costituiscono la flora intestinale).
Numerosi batteri, infine, sono patogeni, cioè causano malattie sia alle piante sia agli animali, uomo compreso (nei cui confronti possono agire attaccando i tessuti direttamente o producendo tossine).
Sistematica
Una relativamente recente proposta di classificazione generale delle monere, che sembra destinata a soppiantare le classificazioni tradizionali (alquanto controverse), è quella che distingue due raggruppamenti: quello molto piccolo degli archeobatteri (o batteri antichi) e quello degli eubatteri (o batteri veri), che riunisce la maggior parte di questi organismi.
Gli archeobatteri( studiati a fondo solo di recente) presentano una serie di caratteristiche peculiari: per esempio, i lipidi della membrana plasmatica, la composizione della parete e l’RNA ribosomico sono differenti da quelli delle altre cellule procarioti e di quelle eucarioti. Sono probabilmente batteri primitivi, che vivono in condizioni oggi considerate estreme, ma che un tempo dovevano essere quelle più comuni sulla Terra.
Fanno parte degli archeobatteri, tra gli altri:
- i metanobatteri, che producono metano riducendo CO2 e H2
- i batteri termoacidofili, che vivono nelle sorgenti termali sulfuree, acide e calde
- i batteri alofili, che vivono in acque salate molto concentrate (come quelle del Mar Morto).
Gli eubatteri riuniscono, come detto, la maggior parte delle classi di batteri conosciute, di cui fanno parte specie di grande utilità per l’uomo, come pure specie patogene.
Particolare importanza tra gli eubatteri rivestono i cianobatteri, capaci, come detto, di fotosintesi.

Schema strutturale di un Methanobrevibacter smithii, in cui si vedono la membrana cellulare (ocra) e la parete cellulare (violetto)
I cianobatteri
I cianobatteri sono batteri aerobi fotosintetici. Sono detti anche cianoficee, o alghe azzurre, perché contengono, oltre alla clorofilla, la ficocianina, pigmento che conferisce il caratteristico colore blu-verde. La parete cellulare è rinforzata da uno strato di mucillagine che tiene uniti gli individui in colonie; la fotosintesi avviene nei tilacoidi, non inseriti in un cloroplasto. Si riproducono agamicamente.
Vivono nelle acque salate o dolci, spesso putride e ricche di detriti organici, che possono utilizzare direttamente.
Nei periodi di particolare crescita o “fioritura” si accumulano in superficie, creando problemi agli impianti idrici e agli altri organismi poiché riducono l’ossigeno disponibile per gli altri organismi; molte specie producono tossine. Si trovano anche nel terreno, sulle cortecce degli alberi o sulle rocce umide vicino a sorgenti termali.
I cianobatteri sono tipici organismi pionieri, capaci anche di fissare l’azoto atmosferico (sono importanti nella produttività delle risaie); alcune specie vivono in simbiosi con ife fungine formando i licheni.
Si pensa che i cianobatteri siano stati tra i primi organismi viventi, insieme ai batteri fotosintetici anaerobi: lo testimonierebbero le stromatoliti, accumuli di calcare depositati dai cianobatteri in ambienti di acqua bassa datati circa 3,5 miliardi di anni fa.

Colonie sferiche di cianobatteri d'acqua dolce, della specie Nostoc pruniforme.
Il regno dei protisti
I protisti comprendono organismi eucarioti unicellulari e pluricellulari con tessuti non differenziati. Comprendono specie che possono essere considerate ai confini tra i regni delle piante, degli animali e dei funghi. Si riproducono per lo più asessualmente, per divisione cellulare, e talvolta sessualmente, per coniugazione. Secondo il modo di procurarsi il nutrimento, i protisti si possono distinguere in tre grandi raggruppamenti: protisti autotrofi, fotosintetici, assimilabili alle piante; protisti eterotrofi o protozoi, assimilabili agli animali; protisti saprofiti, che si nutrono per assorbimento, assimilabili ai funghi.

Composizione di vari protisti
I protisti autotrofi
I protisti autotrofi sono acquatici e comprendono quasi tutte le alghe, unicellulari e pluricellulari. Le alghe unicellulari costituiscono complessivamente la biomassa prevalente del fitoplancton, responsabile di quasi il 70% di tutta l’attività fotosintetica della Terra. A questo gruppo appartengono tre phyla principali: le euglenofite, le crisofite e le pirrofite.
Le euglenofite (dal greco, alghe con la pupilla) si muovono mediante un flagello; una macchia oculare rileva la luce e quando non è sufficiente perdono i cloroplasti e si comportano da eterotrofi. Vivono nelle acque dolci e si riproducono per scissione.
Le crisofite (dal greco, alghe d’oro) possiedono placche calcaree o silicee e, oltre alla clorofilla, un pigmento giallo, la fucoxantina. Si riproducono sia asessualmente, sia sessualmente. Hanno dimensioni minime e costituiscono pertanto il cosiddetto nannoplancton (insieme di organismi acquatici microscopici che galleggiano alla deriva).
Comprendono le diatomee, con guscio siliceo, i cui depositi fossili hanno prodotto la cosiddetta “farina fossile”.
Le pirrofite( dal greco, alghe di fuoco), o dinoflagellati, si muovono mediante due flagelli. Possono avere un guscio di cellulosa diviso in placche da scanalature. Alcune specie, prive di pigmenti, sono eterotrofe. Vivono nell’acqua dolce, ma soprattutto nel mare; molte specie sono luminescenti. Alcuni (Gonyaulax) possiedono pig-menti rossi, che mascherano la clorofilla e sono responsabili del fenomeno della colorazione rossa delle acque. Altri (zooxantelle) vivono in simbiosi con i coralli.

Assortimento di diatomee
Le alghe
Le alghe sono piante acquatiche. Possono essere unicellulari (aggregate in colonie) o pluricellulari, il cui corpo è formato da tessuti indifferenziati (tallo).
Le alghe pluricellulari hanno tuttavia sviluppato dei rizoidi, con i quali si attaccano al substrato.
Le alghe sono le prime piante comparse sulla Terra: le alghe pluricellulari derivano probabilmente da protisti unicellulari autotrofi che non si sono divisi completamente, dando origine a forme filamentose, dapprima semplici, poi ramificate e laminari. Tutte le alghe sono importanti produttori primari degli ambienti acquatici; alcune sono utilizzate anche dall’uomo per diversi scopi.

Caulerpa taxifolia
Riproduzione della alghe
Le alghe si riproducono agamicamente, cioè in modo asessuato, per scissione o mediante spore, e sessualmente, con alternanza di generazioni in cui predomina la fase gametofitica.
Classificazione
La sistematica delle alghe è piuttosto complessa e controversa: molti autori le considerano eucarioti primitivi; altri le includono nel Regno delle Piante.
Le alghe sono comunque classificate secondo il tipo di clorofilla e i pigmenti (xantofille) accessori presenti, che permettono loro di vivere a diverse profondità, dove vi è una differente penetrazione della luce.
Le alghe si distinguono in tre divisioni principali:
- le alghe rosse, o rodofite
- le alghe brune, o feofite
- le alghe verdi, o clorofite
Le alghe rosse, o rodofite, contengono, oltre alla clorofilla, la ficoeritrina, un pigmento che ne maschera il colore verde (conferendo a esse colorazioni dal rosso vivo al nero) e che permette di assorbire le radiazioni luminose verdi-azzurre che penetrano a profondità maggiore. Sono considerate le più primitive per la presenza di pigmenti accessori simili a quelli dei cianobatteri e per la struttura semplice della membrana fotosintetica.
Nelle alghe brune, o feofite, oltre alla clorofilla è presente il pigmento fucoxantina, che ne determina le particolari colorazioni giallo-brune. Il tallo è per lo più frondoso e può raggiungere notevoli dimensioni.
Le alghe verdi, o clorofite, comprendono forme unicellulari flagellate o coccoidi (rotondeggianti e prive di flagelli), isolate o riunite in colonie, che potrebbero aver dato origine alle forme pluricellulari. Contengono, accanto alla clorofilla, diverse xantofille che non ne mascherano tuttavia il colore verde. Sono considerate le più evolute per il tipo di cloroplasti e di materiale di riserva (amido), simili a quelli delle piante superiori.
| DIVISIONE | PIGMENTI | HABITAT | SPECIE |
| rodofite, o alghe rosse | clorofilla
ficoeritrina |
vivono nel mare, soprattutto in quelli più caldi, fino a 170 m di profondità | comprendono le bangioideee le floridee |
| feofite, o alghe brune | clorofilla
fucoxantina |
mari freddi | Fucus ha tallo lungo da 30 cm a 1 m
e cresce sugli scogli tra la bassa e l'alta marea dei mari freddi il sargasso forma dense popolazioni in alto mare; il galleggiamento è favorito da vescicole piene di gas Macrocystis gigantea cresce lungo la costa del Pacifico e raggiunge anche i 100 m di lunghezza Laminaria ha grosse dimensioni, è utilizzata come foraggio e per l'estrazione dello iodio |
| clorofite, o alghe verdi | clorofilla | per lo più di acqua dolce, ma anche marine | clorofite unicellulari sono: Volvox, che forma colonie sferiche in cui alcune cellule si sono specializzate per la riproduzione e vive per lo più nelle acque dolci; Chlamydomonas, flagellata; Chlorella, che produce un antibiotico pluricellulare è Ulva lactuca, detta lattuga di mare per la forma e le dimensioni, comune nel Mediterraneo |

Alghe bianche, rosse e verdi
I protisti eterotrofi, o protozoi
I protisti eterotrofi, o protozoi (dal greco, primi animali) sono prevalentemente acquatici; alcuni sono parassiti, altri sono simbionti, moltissimi sono presenti dove abbondano le sostanze organiche in decomposizione. Comprendono quattro gruppi principali: zooflagellati, sporozoi, sarcodini e ciliati.
Gli zooflagellati sono considerati i protozoi più antichi. Vivono nel suolo e nell’acqua, dove si muovono mediante un flagello. Alcuni sono simbionti o parassiti: Trypanosoma gambiense è l’agente della malattia del sonno, trasmessa dalla mosca tse-tse; altri vivono nell’intestino delle termiti, dove partecipano alla digestione della cellulosa.
I sarcodini possiedono prolungamenti citoplasmatici (pseudopodi) che permettono il movimento (ameboide) e la cattura delle particelle alimentari. Le amebe non hanno forma definita; alcune sono parassite e possono provocare disturbi intestinali nell’uomo (amebiasi). I foraminiferi, marini, hanno gusci calcarei che, accumulandosi sul fondo del mare, formano la sabbia a foraminiferi. I radiolari, marini, hanno invece scheletri silicei che formano la sabbia a radiolari.
Gli sporozoisono tutti parassiti e formano spore. Emettono pseudopodi per catturare il cibo, ma si muovono scivolando o flettendosi. Alternano riproduzione sessuale e asessuale. Passano da un organismo all’altro utilizzando spesso gli insetti come vettori. Plasmodium, trasportato dalla zanzara anofele, provoca la malaria nell’uomo.
I ciliati possiedono caratteristiche ciglia, che possono in alcune specie ricoprire tutto il corpo. Sono diffusi soprattutto nelle acque dolci, in forme libere o fisse a un substrato; sono detti anche infusori perché scoperti negli infusi di fieno. Si riproducono per scissione e per coniugazione. Sono considerati i protisti più evoluti; comprendono i generi Paramecium, Vorticella, Stentor.

Ammonia beccarii, foraminifero bentonico del Mare del Nord.
I protisti saprofiti
I protisti saprofiti comprendono organismi simili a funghi, che si nutrono per assorbimento di sostanze in decomposizione. Sono privi di parete cellulare e vengono chiamati funghi mucillaginosi. Si dividono in due gruppi principali: gli acrasiomiceti e i mixomiceti.
Gli acrasiomiceti hanno struttura cellulare. Sono cellule ameboidi, con prolungamenti citoplasmatici o pseudopodi. In condizioni avverse, possono riunirsi in un denso aggregato in cui alcune cellule si specializzano per compiti diversi.
I mixomiceti non hanno struttura cellulare individuale: formano una massa di citoplasma con miliardi di nuclei non separati da una membrana, che può diffondersi in uno strato molto sottile per alcuni metri quadrati. Di colore giallo-arancio, vivono nelle foglie e nei tronchi marcescenti. Si riproducono per spore.

Fuligo septica
Le piante e i funghi
Al regno delle piante appartengono organismi con una grande varietà di forme, che corrispondono a diversi stadi evolutivi. Comprendono le briofite, forme di transizione dall’ambiente acquatico a quello terrestre, e le piante terrestri più evolute, comprendenti le spermatofite, dotate di semi (che includono le piante con fiore o angiosperme).
Il regno dei funghi comprende organismi probabilmente derivati dalle alghe, che hanno seguito un percorso evolutivo differente da quello delle piante, perdendo la capacità di fotosintesi e divenendo eterotrofi.
Il regno delle piante
Al regno delle piante appartengono gli organismi eucarioti, autotrofi (capaci di fotosintesi), pluricellulari (tranne poche eccezioni).
Le piante considerate più primitive (o piante inferiori) sono le tallofite, così dette perché hanno un corpo costituito da tessuti indifferenziati, detto tallo. Le tallofite acquatiche sono raggruppate sotto il nome di alghe (Protisti) mentre quelle considerate forme di passaggio dall’ambiente acquatico a quello terrestre sono denominate briofite.
Le piante più evolute (o piante superiori) sono le cormofite terrestri, con un corpo differenziato in radici, fusto e foglie, detto cormo; queste piante sono denominate anche piante vascolari, o tracheofite perché dotate di vasi conduttori, o trachee; sono distinte in pteridofite, che comprendono le forme più primitive che si riproducono per spore, e in spermatofite, che comprendono le forme più evolute, perché si riproducono mediante semi.
Tutte le piante mostrano un’alternanza di generazione tra sporofito e gametofito, con una graduale diminuzione della fase gametofitica rispetto a quella sporofitica dalle piante più primitive (alghe) a quelle più evolute (spermatofite).
| tallofite | briofite (terrestri) | ||
| cormofite (tracheofite) | pteridofite (con spore) | psilofite; licofite sfenofite; pterofite | |
| spermatofite (con semi) | gimnosperme (con semi nudi) | cicadofite; coniferofite;
ginkgofite |
|
| angiosperme (o antofite, con semi protetti) | monocotiledoni
dicotiledoni |
Le briofite
Le briofite sono tallofite evolute probabilmente come adattamento di un’alga verde all’ambiente terrestre. Pur non avendo ancora tessuti dfferenziati, in molte briofite si possono distinguere un fusticino, minuscole foglioline e rizoidi per attaccarsi al terreno. Non avendo tessuti di conduzione e di protezione sviluppati per la vita sulla terraferma, mantengono piccole dimensioni (raggiungono un’altezza di pochi centimetri).

Bryum argenteum
Riproduzione delle briofite
Il ciclo vitale delle briofite comprende un’alternanza di generazioni. Nel gametofito aploide dominante i gameti femminili sono prodotti negli archegoni e i gameti maschili negli anteridi. Lo sporofito diploide, molto ridotto, cresce sul gametofito ed è costituito da un peduncolo (seta) e da una capsula nella quale si formano meiospore, dalla cui germinazione deriverà il nuovo gametofito.
La riproduzione delle briofite è ancora strettamente legata all’acqua: i gameti maschili, per poter raggiungere l’archegonio femminile devono nuotare per mezzo di flagelli anche solo in un velo d’acqua; per questo le briofite vivono nel sottobosco, in luoghi molto umidi e ombrosi. Alcune specie possono tuttavia sopravvivere anche dove l’acqua è scarsa, come nei deserti o sulla nuda roccia.

Ciclo vitale delle briofite
Classificazione delle briofite
Le briofite comprendono le epatiche e i muschi.
Le epatiche hanno tallo a forma di lamina e sono frequenti soprattutto nelle regioni di caldo umido. Vivono sul terreno, dove possono formare densi tappeti verdi, ma anche sulla corteccia degli alberi.
I muschi formano densi cuscinetti di minuscoli individui. Insieme ai licheni, i muschi sono organismi pionieri che sgretolano le rocce e rendono possibile la formazione di humus e la crescita di altre piante. Gli sfagni sono muschi di colore chiaro che vivono ai margini degli stagni e dalla cui decomposizione si forma la torba, usata come combustibile e in giardinaggio, per la capacità di trattenere l’acqua e cederla lentamente.

Lunularia cruciata, una epatica
Le tracheofite
Le tracheofite sono le piante che hanno sviluppato tessuti specializzati (conduttori e meccanici) necessari per la vita sulla terraferma.
I tessuti conduttori, (il nome tracheofite deriva da trachee, un tipo di tessuto conduttore), costituiscono un sistema di vasi (per cui le tracheofite sono dette anche piante vascolari) specializzati per il trasporto dell’acqua e dei sali minerali prelevati dal terreno, e la redistribuzione degli zuccheri, formati dalle foglie mediante fotosintesi, a tutto il corpo.
I tessuti meccanici hanno la duplice funzione di sostegno e di protezione contro il disseccamento e sono indispensabili in un ambiente in cui il corpo non è sostenuto dall’acqua.
Le tracheofite hanno sviluppato una struttura organizzata in apparati con funzioni specifiche (cormo), che comprende:
- radice (sistema assorbente e di conduzione)
- fusto (sistema con funzione di sostegno e di conduzione)
- foglie (sistema fotosintetico e traspirante)
Come ulteriore evoluzione, nelle tracheofite lo sporofito predomina sul gametofito. Le tracheofite sono classificate secondo il tipo di riproduzione; si distinguono:
- tracheofite con spore, o pteridofite (licopodi, equiseti e felci)
- tracheofite con seme, o spermatofite, che comprendono le gimnosperme e le angiosperme

La foglia è la parte della pianta in cui avviene la maggiorparte della fotosintesi
Pteridofite
Molto diffuse nel Carbonifero, con organismi di grandi dimensioni che costituivano enormi foreste, sono oggi ridotte a piccole piante del sottobosco.
La riproduzione avviene mediante spore aploidi che formano un piccolo gametofito, in genere laminare (protallo); nel gametofito si formano i gameti maschili e femminili e dopo la fecondazione si sviluppa uno sporofito frondoso.
Come nel caso delle briofite, le pteridofite sono ancora legate all’ambiente acquatico per la riproduzione (i gameti maschili “nuotano” in un velo d’acqua fino ai gameti femminili immobili); inoltre, i piccoli gametofiti non hanno vasi conduttori. Alle pteridofite appartengono le felci.
| licofite | comprendono i licopodi, piccole piante alte pochi centimetri, con foglie a scaglie, come quelle dei muschi |
| sfenofite | comprendono il solo genere Equisetum, che non raggiunge il metro di altezza. Le spore sono prodotte in particolari strutture a cono all'apice dei fusti |
| pterofite | comprendono 12.000 specie di felci, con foglie larghe, dette fronde; sulla pagina inferiore portano le spore riunite in sporangi detti sori. Oggi solo le felci tropicali sono arboree |

Sottobosco di felci, boschi nei dintorni di Cantù
Spermatofite
Negli ultimi 250 milioni di anni le spermatofite hanno invaso tutti gli ambienti terrestri, diventando le piante dominanti grazie a una più efficace strategia riproduttiva basata su due adattamenti principali: il polline e i semi.
Il polline è un insieme di microscopici granuli (da 2,5 a 250 micron) contenenti cellule riproduttive, i gameti maschili. Costituisce dunque il gametofito maschile (ridotto quindi a poche cellule), mentre lo sporofito è la pianta, spesso di notevoli dimensioni.
Ogni granulo di polline è rotondeggiante (in genere di colore giallo-arancio), formato da due cellule protette da un involucro esterno, rigido, con rilievi e disegni caratteristici per ogni specie.
Vengono prodotte grandi quantità di polline che, leggero e minuto, viene trasportato facilmente dal vento o dagli animali e garantisce una sicura diffusione fino a raggiungere il gametofito femminile (impollinazione): con questo accorgimento la fecondazione è completamente svincolata dall’acqua.
Il seme è una struttura protettiva che avvolge con un rivestimento rigido l’embrione e una riserva alimentare per la sua crescita. Il seme permette di mantenere l’embrione in uno stato di quiescenza finché non vi siano le condizioni ottimali per la germinazione.
Per assicurarne la dispersione, i semi possiedono caratteristiche che li rendono appetibili agli animali, o facilmente trasportabili dall’acqua e dal vento per grandi distanze; in questo modo le spermatofite hanno potuto diffondersi rapidamente.
Le spermatofite comprendono le gimnosperme (spermatofite con seme nudo) e le angiosperme (spermatofite con fiori).

Girasole (Helianthus annuus)
Le gimnosperme
Le gimnosperme sono spermatofite con semi “nudi”, cioè non avvolti da strutture di protezione.
Gli organi della riproduzione sono ciuffi di foglie trasformate o sporofilli (squame) riuniti intorno a un asse centrale (rachide) in grappoli unisessuali detti strobili, o coni: quelli maschili (microsporofilli), in genere più piccoli e situati all’estremità dei rami più bassi, portano le sacche polliniche, in cui si svilupperanno i gameti maschili (cellule spermatiche); quelli femminili (macrosporofilli), più grandi, si trovano sui rami più alti e portano gli ovuli, in cui si svilupperanno i gameti femminili (cellule uovo).
L’impollinazione è esclusivamente anemofila: i coni maschili liberano immense nuvole di polline che vengono trasportate dal vento. Alla base di ogni squama dei coni femminili si formano due ovuli, contenenti ciascuno una cellula uovo che viene fecondata dalla cellula spermatica di un granulo di polline atterrato vicino.
La cellula uovo fecondata rimane racchiusa nell’ovulo, che diventa una struttura rigida, il seme, in cui si sviluppa l’embrione. Nel frattempo i macrosporofilli lignificano e si aprono a maturità liberando i semi.
Le gimnosperme sono tutte piante legnose e si dividono in ginkgofite, cicadofite e coniferofite.
| ginkgofite | molto diffuse dal Giurassico, oggi sono quasi scomparse e considerate fossili viventi. L'unica vivente, Ginkgo biloba, diffusa nei parchi e nei giardini, ha foglie a ventaglio incise a metà |
| cicadofite | assomigliano a palme, con un ciuffo di foglie all'estremità di un tronco non ramificato; vivono nei paesi tropicali; le radici ospitano microrganismi azotofissatori |
| coniferofite | sono così chiamate perché portano i coni, o pigne; hanno tutte foglie aghiformi, cioè sottili e allungate, in genere appuntite all'apice, come adattamento alla mancanza di acqua: vivono infatti in alta montagna dove per le basse temperature invernali l'acqua gela e non è disponibile per diversi mesi, e nelle regioni mediterranee con scarsa piovosità. Possiedono nella linfa una sostanza che agisce come antigelo, la resina, che permette il trasporto delle sostanze nutritive anche con temperature sotto lo zero. Comprendono i pini, gli abeti, il larice, il cipresso, il tasso, il ginepro, tutte sempreverdi tranne il larice: mantenere le foglie tutto l'anno permette di risparmiare in primavera acqua ed energia per formare i nuovi germogli |

Pigne maturande di Abete del Colorado (Picea pungens)
Le angiosperme
Le angiosperme, dette anche magnoliofite, sono piante a seme provviste di fiore, con ovuli protetti in un involucro chiuso (ovario), che in seguito a fecondazione si sviluppa nel frutto(angiosperma significa infatti “seme racchiuso”).
Le angiosperme costituiscono oggi il gruppo dominante di piante: comprendono circa 230.000 specie, diffuse in ogni ambiente (piante acquatiche, terrestri, parassite). Il loro successo è dovuto a diversi adattamenti, i principali dei quali sono il fiore e il frutto.
Il fiore è una struttura riproduttiva complessa, derivata dalla trasformazione di foglie, in cui si formano e sono protetti i gametofiti maschili e femminili, che, come nelle gimnosperme, sono ridotti a poche cellule del granulo pollinico e dell’ovulo.
Il fiore, spesso con petali colorati, ha una funzione vessillare, rivolta cioè ad attirare gli insetti o gli altri animali impollinatori, garantendo così un maggiore successo riproduttivo.
Anche la fecondazione avviene nel fiore e lo zigote rimane racchiuso nell’ovulo trasformato in seme.
Il frutto è una struttura di protezione del seme derivata dalla trasformazione del fiore, dopo la fecondazione. In genere è colorato e dolce per essere appetito dagli animali, o dotato di adattamenti per essere trasportato dal vento (in modo da assicurare una più ampia dispersione del seme).
Il successo delle angiosperme è stato favorito anche dalle foglie a lamina larga (latifoglie), che assorbono maggiormente la luce per una più efficace fotosintesi.
La sistematica delle angiosperme si basa sulle caratteristiche del fiore, e primariamente sulla presenza nel seme di una o due foglioline embrionali (cotiledoni): si dividono così in monocotiledoni(un solo cotiledone) e dicotiledoni (due cotiledoni).
Le più importanti monocotiledoni sono le glumiflore (a cui appartengono i cereali, fondamentali nell’alimentazione umana), specie alimentari come il banano e le palme, specie conosciute per i fiori come le liliacee e le orchidee.
Le dicotiledoni sono più diffuse: a esse appartengono le principali piante da frutto e innumerevoli specie arboree, arbustive ed erbacee.

Fiore del genere Magnolia
Il regno dei funghi
I funghi sono organismi eterotrofi, in genere con corpo filamentoso pluricellulare indifferenziato (tallo), che si nutrono per assorbimento.
Derivano probabilmente, come le piante, dalle alghe, ma presto nel corso dell’evoluzione hanno perso il carattere distintivo delle piante, cioè l’autotrofia.

Composizione di Fungi
Struttura e riproduzione
Le cellule dei funghi hanno per lo più una parete cellulare di micosina, una sostanza azotata simile alla chitina degli artropodi.
Il tallo è formato da una massa di sottili ife filamentose detta micelio, che si accresce nel terreno. In alcune specie le ife sono singole cellule allungate con più nuclei, in altre possono essere formate da più cellule divise da setti, perforati per il passaggio delle sostanze nutritive.
Poiché hanno una parete rigida, i funghi si nutrono per assorbimento, secernendo all’esterno enzimi digestivi e assorbendo le sostanze nutritive che vengono così liberate. Le ife si estendono per una grande superficie coprendo così una vasta area di assorbimento.
Nei funghi si osservano diversi tipi di riproduzione, spesso complessi e generalmente con dominanza della generazione aploide.
La riproduzione asessuata avviene per frammentazione del micelio. Periodicamente, le ife si aggregano in strutture riproduttive, dette corpi fruttiferi, che sono le sole strutture visibili del fungo (quelle che si mangiano nei funghi commestibili, per esempio). Nei corpi fruttiferi si sviluppano, per via sia sessuata sia asessuata, microscopiche spore, che formano un polvere scura facilmente dispersa dal vento.

Riproduzione sessuata dei funghi
Importanza ecologica
I funghi vivono in ambienti umidi del sottobosco e sono, con i batteri, importanti decompositori; infatti la maggior parte dei funghi è saprofita (si nutre cioè di materia organica in decomposizione), e molti sono parassiti. Alcuni funghi formano simbiosi mutualistiche con le piante, come nel caso dei licheni, un’associazione tra un fungo e un’alga, e delle micorrize, che invadono le cellule radicali di molte piante vascolari, con cui dividono le sostanze nutritive.

Mycena galericulata (basidiomiceti)
Classificazione dei funghi
Anche la sistematica dei funghi è complessa e soggetta a continue revisioni. Generalmente i funghi sono suddivisi in cinque principali gruppi:
- zigomiceti
- oomiceti
- ascomiceti
- basidiomiceti
- funghi imperfetti o deuteromiceti
Gli zigomiceti formano spore sessuali, dette zigospore, per unione di due ife aploidi. Comprendono il fungo coprofilo (che ha cioè il suo habitat nelle feci) Pilobolus e la muffa nera del pane Rhizopus.
Negli ascomiceti le spore sono disposte in strutture a sacco, dette aschi, che si aggregano a formare corpi fruttiferi complessi, l’ascocarpo. Gli ascocarpi possono avere forme diverse, sulle quali si basa la sistematica della divisione. I più primitivi sono i cleistoteci, sferici; il peritecio ha invece forma a fiasco aperto; più evoluta sarebbe la forma ad apotecio, simile a una scodella aperta.
Gli ascomiceti possono riprodursi sessualmente con la produzione, in seguito a meiosi, di ascospore, o asessualmente con la produzione di conidi, speciali spore portate all’estremità di ife. Durante i periodi sfavorevoli gli ascomiceti possono produrre anche spore durature (clamidiospore), con pareti spesse e ricche di sostanze di riserva.
Gli ascomiceti comprendono molte muffe che si sviluppano sui cibi avariati o sui vegetali e il ricercato tartufo, che cresce completamente sotto terra. Ascomiceti unicellulari sono i lieviti, microrganismi capaci di fermentazione alcolica: in scarsità di ossigeno trasformano gli zuccheri in alcol, e questo processo è sfruttato per la produzione del pane, della birra, del vino. Saccharomyces si riproduce per gemmazione ma può formare ascospore.
Nei basidiomiceti le spore sono contenute in strutture a forma di clava, i basidi, portate spesso su sottili filamenti detti sterigmi. Il corpo fruttifero ha per lo più la tipica forma a cappello sorretto da un gambo. Sulla parte inferiore del cappello i basidi e le ife sterili formano uno strato continuo, o imenio, che riveste dei tubuli o delle lamelle. Alcuni basidiomiceti hanno corpo fruttifero crostoso e a mensola (come i funghi che crescono sui tronchi degli alberi); altri, considerati i meno evoluti, non hanno corpo fruttifero e sono per lo più parassiti delle piante.
I basidiomiceti sono considerati i funghi più evoluti: nel loro ciclo di sviluppo domina la generazione diploide. Comprendono il maggior numero dei funghi conosciuti, con specie commestibili, velenose e parassite.
I funghi imperfetti comprendono specie diverse, che apparentemente mancano di riproduzione sessuale. Tra queste ricordiamo Penicillium, da cui si ottiene la penicillina, alcune specie responsabili di malattie (micosi) come la tricofitosi e il piede d’atleta e alcuni funghi predatori dei vermi nematodi.
Gli oomiceti hanno pareti cellulari che spesso contengono cellulosa invece di chitina. Nella riproduzione sessuale producono grandi cellule uovo, in quella asessuale producono, unici tra i funghi, zoospore mobili nell’acqua. Comprendono molti funghi responsabili di malattie delle piante come la peronospora della vite e della patata e le muffe d’acqua che vivono nel terreno umido o nell’acqua.

zigomicete |

Tartufo, Tuber (ascomiceti) |

Penicillium su alcuni mandarini |

Peronospora su un grappolo di pomodori, Phytophthora infestans (oomicete) |
I licheni
I licheni sono associazioni vegetali formate da funghi e alghe, che vivono in stretta simbiosi formando un unico tallo. Il micelio dei funghi, per lo più ascomiceti, assume acqua e sali minerali dal substrato e trattiene l’umidità; le alghe, clorofite unicellulari o cianobatteri, producono gli zuccheri.
I pigmenti delle alghe sono responsabili dei colori vivaci di alcuni licheni, dal verde all’arancio.
La riproduzione avviene per:
- diffusione delle spore dei funghi, che, se incontrano un’alga adatta, potranno ricostituire la struttura del lichene
- per frammentazione del tallo (tali frammenti sono detti isidi e soredi).
La crescita è lentissima.
I licheni si dividono secondo la forma del tallo in licheni crostosi, con tallo sottile attaccato completamente al substrato; fogliosi, con tallo orizzontale simile a una foglia; fruticosi, a forma eretta o cespugliosa, che crescono direttamente nel terreno o pendono dai rami.
I licheni sono i primi organismi pionieri che colonizzano le rocce e le sgretolano secernendo degli acidi: si inizia così il processo di formazione del suolo.
I licheni crescono in qualunque ambiente con molta luce, sopportando lunghi periodi di clima secco.

Cladonia portentosa lichene composto
Il regno degli animali
Il regno degli animali comprende gli organismi pluricellulari eterotrofi che si riproducono sessualmente e le cui cellule, a differenza di quelle vegetali, sono prive di parete cellulare. Derivati presumibilmente da protisti che formavano colonie marine simili alle attuali spugne, gli animali si sono eveluti in una straordinaria varietà di forme, con livelli di complessità i più diversificati, connessi a una crescente organizzazione e specializzazione cellulare in tessuti. I phyla animali riflettono questa tendenza a una sempre maggiore complessità.

Composita di Animalia
Tendenze evolutive degli animali
Sono animali gli organismi che, pur nella loro varietà, hanno in comune le seguenti caratteristiche principali:
- sono pluricellulari
- sono eterotrofi
- hanno cellule prive di parete cellulare
- si riproducono sessualmente (in molte specie, tuttavia, la riproduzione è anche agamica)
- sono mobili, almeno in un periodo della loro vita
- reagiscono agli stimoli esterni
Come le piante, anche gli animali hanno subìto un processo di evoluzione che ha portato a un’organizzazione cellulare sempre più complessa. Questa tendenza alla complessità si osserva nelle seguenti caratteristiche:
- organizzazione cellulare
- disposizione simmetrica degli organi e segmentazione
- cefalizzazioneevoluzione di una cavità interna
- segmentazione
- evoluzione di strutture di sostegno
- evoluzione del canale digerente
Il grado via via crescente di complessità delle strutture e delle funzioni svolte da questi organismi costituisce la chiave per classificare gli animali.
I prossimi paragrafi descrivono le tendenze evolutive degli animali. Segue una breve descrizione dei principali phyla in cui gli animali vengono classificati.
| poriferi | calcisponge
ialosponge demosponge |
| cnidari | idrozoi
scifozoi antozoi |
| ctenofori | |
| tentacolati | turbellari
trematodi cestodi |
| platelminti | |
| gnatostomulidi | |
| nemertini | |
| aschelminti | |
| sipunculidi | |
| camptozoi | |
| anellidi | policheti
oligocheti irudinei |
| pentastomidi | |
| tardigradi | |
| onicofori | |
| artropodi | aracnidi
crostacei miriapodi insetti |
| molluschi | gasteropodi
scafoidi lamellibranchi cefalopodi |
| echinodermi | crinoidei
asteroidei ofiuroidei echinoidei oloturoidei |
| cordati | cefalocordati
urocordati
vertebrati: |
Schema di classificazione degli animali
Organizzazione cellulare
Nel corso dell’evoluzione le cellule degli animali si specializzano e si organizzano in tessuti, i quali a loro volta si uniscono a formare gli organi. Anche gli organi si uniscono in apparati o sistemi di organi.
Si osserva una tendenza alla complessità anche per quanto riguarda il numero dei foglietti embrionari (strati di tessuto presenti nell’embrione): accanto ai due strati iniziali (ectoderma ed endoderma), si forma in uno stadio ulteriore, un terzo strato intermedio (mesoderma).

Gastrulazione di un diploblastico: La formazione dei foglietti germinativi (o embrionali) da (1) blastula a (2) gastrula. Alcune cellule dell'ectoderma (in arancione) si introflettono a formare l'endoderma (in rosso).
Simmetria e segmentazione
Gli animali più semplici (le spugne) sono privi di simmetria.
Il primo tipo di disposizione simmetrica degli organi è a simmetria raggiata: gli organi sono disposti attorno a un asse centrale e il corpo dell’animale può essere idealmente attraversato da infiniti piani passanti per l’asse, che lo dividono in due parti uguali. Questo tipo di simmetria è caratteristico degli animali acquatici meno evoluti, che vivono fissi a un substrato (animali sessili) o sono poco mobili.
Propria degli animali più evoluti e capaci di movimento è la simmetria bilaterale, in cui esiste un solo piano che divide il corpo in due parti simmetriche. In questi animali sono presenti una superficie dorsale e una ventrale, un’estremità cefalica e una caudale. In alcuni animali il corpo è segmentato in unità strutturali simili (metameri), in cui gli organi si ripetono con identica disposizione. Negli organismi più complessi la segmentazione non è visibile esternamente, ma riguarda solo alcune parti dei muscoli, dello scheletro e di nervi.

Simmetria radiale della stella marina
Cefalizzazione
La cefalizzazione è il fenomeno per cui gli organi di senso, le cellule nervose e gli organi deputati all’ingestione del cibo si sono localizzati (cioè concentrati) a un’estremità del corpo, cioè la testa dell’animale (estremità cefalica).

Pseudobiceros bedfordi, platelminta (verme piatto), uno degli animali più semplici in cui s'è appena accennata la cefalizzazione
Cavità corporea
Si chiama celoma la cavità interna nella quale sono situati gli organi, interposta tra il canale digerente e la parete interna del corpo. Grazie al celoma, organi quali il cuore, i polmoni, lo stomaco e l’intestino hanno a disposizione un certo spazio in cui accrescersi, muoversi e funzionare senza reciproche interferenze.
Gli animali più semplici sono acelomati, cioè privi di celoma. Altri possiedono uno spazio analogo al celoma (pseudoceloma), la cui origine nella fase embrionale è diversa rispetto agli animali dotati di celoma.
Canale alimentare
Il canale alimentare degli animali più primitivi è un sacco provvisto di una sola apertura, attraverso cui il cibo viene introdotto e i prodotti di scarto espulsi.
Segno di maggiore complessità evolutiva è la presenza di un canale alimentare con due aperture, la bocca e l’ano.
Strutture di sostegno
Si usa distinguere gli animali in vertebrati, provvisti di endoscheletro, cioè di uno scheletro interno o colonna vertebrale, e invertebrati, che ne sono privi e sopperiscono alla funzione di sostegno con diversi accorgimenti: “aghi” rigidi di sostanze minerali, idroscheletri e scheletri esterni, o esoscheletri, di varia natura. I vertebrati costituiscono solo una parte di un singolo phylum, quello dei cordati (che comprende: pesci, anfibi, rettili, uccelli e mammiferi).
| CLASSE | CARATTERISTICHE |
| calcisponge | spugne calcaree; vivono nel mare fino a circa 800m di profondità |
| ialosponge | spugne silicee; vivono nel mare tra 25m e 8500m |
| demosponge | spugne cornee; vivono anche in acque dolci, ma sono più diffuse nel mare. Il loro scheletro di spongina viene utilizzato come spugna da bagno |

Anfiosso (Branchiostoma lanceolatum), il più semplice dei Cordati
Poriferi
I poriferi, o spugne (phylum Poriphera), sono gli animali più semplici, poiché non possiedono né simmetria né veri e propri tessuti. Possono essere considerati delle colonie di organismi unicellulari con funzioni indipendenti.
Le spugne vivono in acqua e hanno il corpo a forma di sacco cavo, fisso al substrato e perforato da numerosi pori, attraverso i quali entrano le particelle alimentari portate dall’acqua. L’interno della cavità è rivestito da particolari cellule (coanociti), provviste di un flagello che, muovendosi, assicura il continuo flusso dell’acqua. Una sorta di “collare” che circonda il flagello funziona da setaccio che trattiene i microrganismi entrati dai pori. I prodotti di rifiuto fuoriescono da un’ampia apertura, l’osculo.
Tra le cellule epiteliali e i coanociti si trova uno strato gelatinoso (mesoglea) che contiene cellule ameboidi, le quali secernono spicole calcaree, silicee o cornee che danno sostegno all’animale.
La riproduzione avviene per gemmazione oppure è di tipo sessuato. Nel primo caso si forma una protuberanza, o gemma, sul corpo della spugna; accrescendosi, la gemma può rimanere attaccata al corpo del genitore oppure staccarsi. Se la riproduzione è sessuata, dalla fecondazione della cellula uovo si forma un larva ciliata libera, che viene dispersa dalla corrente e poi si fissa sul fondo per dare origine a un nuovo individuo.

Una spugna marina (Aplysina aerophoba)
Cnidari, o celenterati
Gli cnidari, o celenterati (phylum Cnidaria), sono invertebrati marini dal corpo a forma di polipo e medusa: il polipo è sessile, cioè vive fisso al substrato, solitario o riunito in colonie; la medusa viene trasportata liberamente dalle correnti.
Polipo e medusa rappresentano due fasi del ciclo vitale del celenterato: dalla fecondazione della cellula uovo, liberata da una medusa, si forma una larva ciliata, liberamente natante, che dopo un po’ di tempo si fissa al substrato, trasformandosi in polipo. Questo, a sua volta, si allunga e si divide trasversalmente in numerose parti provviste di tentacoli e impilate l’una sull’altra. Staccandosi, ognuna di esse forma una piccola medusa (efira), che si accresce fino a diventare medusa sessualmente matura. A questo punto il ciclo ricomincia.
Il corpo dei celenterati, a simmetria raggiata, è costituito in larga parte da acqua e ha perciò aspetto gelatinoso. Sono presenti due strati cellulari (ectoderma ed endoderma) che circondano la cavità gastrovascolare. Questa è tappezzata da numerose cellule ghiandolari con funzioni digestive ed è comunicante con l’esterno mediante un’apertura circondata da tentacoli. Sulla superficie dei tentacoli sono distribuiti gli cnidoblasti, particolari cellule che, toccate da un corpo estraneo, estroflettono un filamento cavo. Quest’ultimo inocula nell’aggressore o nella preda un liquido urticante e paralizzante.
Cellule nervose con lunghi prolungamenti (cellule neuroepiteliali) costituiscono un rudimentale sistema nervoso, che consente all’animale di reagire velocemente a uno stimolo ambientale. Specie tipiche sono gli anemoni di mare, le madrepore e i coralli.
| CLASSE | CARATTERISTICHE |
| idrozoi | la generazione di polipo e di medusa si alternano |
| scifozoi | prevale la generazione di medusa |
| antozoi | hanno solo la forma di polipo; comprendono le attinie e i coralli. Secernono uno scheletro calcareo che nel corso di milioni di anni ha formato grandi barriere coralline |

Anemone di mare (Actinaria)
Ctenofori
Gli ctenofori (phylum Ctenophora) sono anch’essi animali marini, i primi in cui allo stadio larvale compare il terzo foglietto embrionario, il mesoderma.
Come i celenterati, anche gli ctenofori hanno il corpo gelatinoso e trasparente, formato per il 95% da acqua. Possiedono una bocca e, in posizione opposta, un organo di senso statico (organo apicale) dal quale si dipartono 8 strisce ciliate, chiamate palette vibratili, utilizzate per la locomozione. Certe specie sono provviste di due lunghi tentacoli che possono secernere sostanze adesive.
Sono animali ermafroditi, cioè uno stesso individuo produce gameti sia maschili sia femminili.

Ctenoforo tentacolato
Tentacolati
I tentacolati (phylum Tentaculata) sono piccoli animali marini, che vivono isolati o in colonie. Il nome del phylum deriva da una corona di tentacoli (lofoforo) che circonda la bocca; il lofoforo crea un vortice d’acqua che convoglia gli alimenti verso la bocca.
Alcuni tentacolati sono protetti da una conchiglia bivalve, altri vivono in tubuli, altri sono vermiformi.
Gli individui sono ermafroditi e la riproduzione può essere sia agamica sia sessuale.
Comprendono i foronoidei, i briozoi e i brachiopodi. Tra questi vanno segnalati i briozoi, che sono sessili e vivono in colonie; hanno inizialmente simmetria bilaterale, ma durante lo sviluppo il corpo si ripiega a U. In questo modo, l’ano e la bocca sono affiancati e rivolti entrambi verso l’alto.
Bathocyroe fosteri
Platelminti
I platelminti (phylum Plathelmynthes) hanno simmetria bilaterale e il corpo piatto (per questo sono anche detti vermi piatti), lungo da pochi millimetri a diversi metri. Sono gli animali più semplici in cui i tessuti siano raggruppati in unità funzionali, gli organi.
La testa ha due macchie oculari, sensibili agli stimoli luminosi ma incapaci di formare immagini. Sono anche presenti aggregati di cellule nervose (gangli) che costituiscono un primitivo cervello. Il sistema nervoso si prolunga per l’intera lunghezza del corpo con 2 cordoni longitudinali uniti da fibre trasversali.
L’apparato digerente è provvisto di una bocca e di una cavità gastrovascolare molto ramificata e a fondo cieco. L’apparato escretore è costituito da tubuli anch’essi a fondo cieco (protonefridi) che terminano con le cellule a fiamma, così chiamate perché provviste di ciglia vibratili, che ondeggiando come una fiammella espellono i prodotti di rifiuto all’esterno del corpo.
La riproduzione può essere sessuale oppure agamica; alcune specie possiedono la capacità di rigenerare parti del loro corpo.
I platelminti comprendono animali a vita libera, marini o terrestri, e parassiti . Questi ultimi (tra cui la tenia, o verme solitario) hanno sviluppato alcuni adattamenti: per esempio, il corpo rivestito da una cuticola a protezione dagli enzimi digestivi dell’ospite; strutture specializzate (ventose e uncini) per afferrarsi alla parete intestinale dell’animale che parassitano; l’ermafroditismo per autofecondarsi.
| CLASSE | CARATTERISTICHE |
| turbellari | conducono vita libera; comprendono la planaria, largamente utilizzata nei laboratori per studiarne le capacità rigenerative |
| trematodi | forme parassite. Comprendono lo Schistosoma (comune in Africa, Asia e Sudamerica, provoca una forma di anemia, nota come schistosomiasi) e la Fasciola hepatica |
| cestodi | tutti parassiti; tra questi, la tenia, dal corpo nastriforme, che vive nell'intestino dell'uomo e di altri animali |

Prostheceraeus giesbrechtii, un verme piatto
Nemertini
I nemertini (phylum Nemertini) sono animali vermiformi, lunghi fino a 30 m, che vivono per lo più sui fondali fangosi dei mari. Sono gli animali più semplici, in cui il canale alimentare è provvisto di un’apertura posteriore, l’ano.
Possiedono inoltre un rudimenatale pseudoceloma (cavità interna) e una specie di proboscide estroflessibile; quest’ultima può essere collegata a ghiandole velenifere ed è impiegata per catturare le prede e, quando non serve, viene riposta in una cavità allungata (rincocele) che decorre dorsalmente al canale alimentare.

Micrura verrilli
Aschelminti
Gli animali che appartengono agli aschelminti (phylum Aschelmintes) hanno caratteristiche molto varie, tanto che alcuni studiosi contestano la parentela tra le diverse classi. Gli elementi che accomunano gli aschelminti sono: simmetria bilaterale, pseudoceloma, sessi separati.
| CLASSE | CARATTERISTICHE |
| rotiferi | sono così chiamati perché possiedono un disco ciliato intorno alla bocca (apparato rotatorio) che, facendo ondeggiare le ciglia, crea un vortice nell'acqua che trascina batteri e protozoi verso la bocca. La loro faringe ha uno spesso strato muscolare che muove dei pezzi scheletrici per sminuzzare il cibo. La riproduzione è sessuale o agamica, ma si riproducono anche per partenogenesi |
| acantocefali | parassiti, sono privi di intestino e di bocca e provvisti all'estremità anteriore di una proboscide armata di uncini |
| gastrotrichi | il nome deriva dalla presenza, in certe specie, di ciglia sulla faccia ventrale. Sono lunghi fino a 1,5 m |
| nematodi | hanno corpo lungo, sottile e cilindrico (sono anche detti vermi cilindrici) rivestito da una cuticola. Lo pseudoceloma ripieno di liquido circonda gli organi e forma uno scheletro idrostatico, contro cui i 4 muscoli longitudinali possono fare forza. La riproduzione è sessuata, generalmente con sessi separati. I maschi si distinguono dalle femmine per la forma e le dimensioni (dimorfismo sessuale).
I nematodi comprendono forme libere, che vivono in tutti gli ambienti, ma sono soprattutto parassiti dell'uomo, degli animali domestici e delle piante |
| nematomorfi | simili ai nematodi, allo stadio larvale sono parassiti |
| chinorinchi | hanno una cuticola provvista di setole o aculei; vivono in mare |
| priapulidi | sono provvisti di una proboscide armata di uncini; vivono nei mari freddi |

Priapulus caudatus, pescato nel mare di Barens
Anellidi
Negli anellidi (phylum Anellida) compare il celoma, ripieno di liquido, con funzione di scheletro idrostatico.
Gli anellidi sono anche detti vermi segmentati, perché il loro corpo è diviso in segmenti, o metameri. In ogni metamero sono presenti due gangli nervosi, una coppia di organi escretori e due vasi sanguigni. I gangli nervosi di ciascun metamero sono uniti tra loro da un cordone trasversale e ai gangli dei metameri adiacenti da un cordone longitudinale.
L’apparato digerente ha inizio con la bocca, situata nel secondo metamero, e termina con l’ano, nell’ultimo segmento.
Compare per la prima volta l’apparato circolatorio, formato da 5 paia di “cuori” che si contraggono ritmicamente e riversano il sangue in due vasi, uno dorsale e uno ventrale; questi ultimi sono uniti, nei singoli metameri, da vasi trasversali.
La respirazione avviene per via cutanea, ma in alcune specie compare un rudimentale apparato respiratorio, rappresentato da espansioni più o meno ramificate (branchie).
L’epidermide dei metameri locomotori può presentare delle espansioni (parapodi) che portano delle setole.
La riproduzione è sessuata e lo sviluppo passa attraverso la larva trocofora, assai interessante dal punto di vista filogenetico, perché simile alla larva dei molluschi.
| CLASSE | CARATTERISTICHE |
| policheti | il corpo è ricoperto da molte setole. Sono diffusi nel mare, liberamente natanti oppure sessili; in questo caso vivono dentro un tubo secreto dalla loro epidermide |
| oligocheti | con poche setole, vivono nelle acque marine, nelle acque dolci e sulla terraferma. Comprendono il lombrico |
| irudinei | privi di setole, sono carnivori o parassiti (per esempio, la sanguisuga) diffusi nelle acque dolci stagnanti e sulla terraferma |

Glycera sp.
Artropodi
Gli artropodi (phylum Arthropoda) sono il phylum più vasto del regno animale. Sono provvisti di appendici articolate atte alla locomozione (da cui il nome del phylum) e di un esoscheletro con funzione protettiva, costituito da chitina e sali di calcio; all’interno dell’esoscheletro si inseriscono i muscoli. Essendo rigido, l’esoscheletro non può accrescersi insieme all’animale e quindi deve essere periodicamente sostituito con un processo chiamato muta.
Gli artropodi hanno un apparato circolatorio aperto (il sangue non scorre nei vasi ma bagna direttamente gli organi interni). Respirano per mezzo di branchie se acquatici, oppure mediante trachee o polmoni a libro se terrestri. L’escrezione avviene grazie a nefridi o tubuli malpighiani che si aprono all’esterno.
I metameri sono riuniti a formare tre regioni distinte: il capo, dove si trovano le antenne e gli occhi composti, il torace con gli arti e l’addome.
Lo sviluppo dell’uovo fecondato è indiretto e avviene attraverso più stadi larvali (metamorfosi).
| CLASSE | CARATTERISTICHE |
| aracnidi | corpo diviso in cefalotorace e addome; cheliceri e pedipalpi per la cattura degli alimenti; quattro paia di zampe; respirazione per trachee o con un tipo particolare di polmoni; specie marine e terrestri |
| crostacei | corpo diviso in cefalotorace (coperto dal carapace) e addome; potenti mandibole; respirazione per branchie; specie per lo più acquatiche |
| miriapodi | corpo segmentato in molti metameri e una testa; apparato escretore costituito da un sistema di tubuli malpighiani; respirazione per trachee; specie terrestri |
| insetti | corpo diviso in 20 segmenti, riuniti in capo, torace e addome; tre paia di zampe e 2 o 4 ali; specie terrestri |

Composita di artropodi
Molluschi
I molluschi (phylum Mollusca) hanno questa particolarità: la loro larva presenta somiglianze con la trocofora degli anellidi e ciò ha fatto avanzare l’ipotesi di un antenato comune.
Tra i due phyla si osservano perciò dei caratteri comuni, ma anche molte differenze.
Il corpo dei molluschi non è diviso in metameri, ma in un capo e un tronco. Il capo ha la bocca, i gangli cerebrali e i principali organi di senso. Il tronco è formato da un muscolo ventrale, chiamato piede, che serve per la locomozione; è presente anche un sacco dei visceri, contenente gli organi, rivestito da una piega cutanea, chiamata mantello; il mantello secerne la conchiglia calcarea.
Nella bocca si trova la radula, una struttura nastriforme provvista di dentelli che servono per triturare il cibo. L’apparato digerente continua con la faringe, l’esofago, lo stomaco e l’intestino; una ghiandola, l’epatopancreas, ha funzione digestiva.
L’apparato circolatorio è aperto (il sangue non scorre nei vasi, ma bagna direttamente i tessuti) e composto da un cuore. I molluschi acquatici respirano per mezzo di branchie a forma di pettine (ctenidi), mentre quelli terrestri attraverso un “polmone” derivato da una porzione della cavità del mantello.
La riproduzione è sempre sessuata (in qualche specie gli individui sono ermafroditi). Lo sviluppo in alcuni casi è diretto, per cui dall’uovo fecondato nasce un individuo simile all’adulto; in altri casi è indiretto, con la formazione di una larva e metamorfosi complessa.
| CLASSE | CARATTERISTICHE |
| bivalvi,
o lamellibranchi, o pelecipodi |
conchiglia a due valve; assenza di un capo definito; acquatici; filtrano il cibo dall'acqua. Comprendono ostriche e mitili |
| cefalopodi | simmetria bilaterale con capo differenziato, provvisto di tentacoli con ventose; mantello saldato, con conchiglia interna, quando presente; sistema nervoso molto sviluppato; marini; un imbuto muscoloso spinge l'acqua permettendo un brusco movimento all'indietro; il nuoto in avanti avviene per mezzo di pinne. Comprendono polpi, seppie, calamari |
| gasteropodi | conchiglia a una sola valva, a spirale; capo ben differenziato, con due occhi spesso in cima a un paio di tentacoli; spesso ermafroditi; presenti in ogni habitat. Comprendono lumache e chiocciole |
| scafopodi | conchiglia a zanna di elefante; vivono sul fondo marino. Comprendono il genere Dentalium |
Polpo comune, Octopus vulgaris
Altri phyla di animali
I gnatostomulidi (phylum Gnathostomulida) comprendono animali di piccole dimensioni (0,5-3 mm), dal corpo cilindrico e vermiforme, ricoperto da ciglia; la loro bocca si apre sul ventre. I gnatostomulidi sono animali ermafroditi e vivono nei fondali sabbiosi e fangosi.
I sipunculidi (phylum Sipunculida) hanno il corpo allungato che si restringe all’estremità anteriore, dove forma una proboscide. Vivono sepolti nei fondali.
I camptozoi, o endoprotti (phylum Kamptozoa), sono animali acquatici che vivono in colonie. Ogni individuo è sostenuto da un peduncolo che in cima si dilata per contenere gli organi. La bocca è circondata da una corona di tentacoli.
I pentastomidi (phylum Pentastomidi) comprendono animali tutti parassiti di vertebrati terrestri. Come adattamento, nello stadio larvale possiedono quattro paia di “arti” muniti di uncini. Durante lo sviluppo, due paia di “arti” scompaiono e le rimanenti due sono riposte in quattro tasche situate attorno alla bocca (da cui il nome del phylum, “cinque bocche”).
I tardigradi (phylum Tardigrada) sono animali di dimensioni microscopiche, con quattro paia di “arti” che terminano con unghie. La bocca è provvista di stiletti retrattili atti a pungere.
Gli onicofori (phylum Onicophora) ricordano gli anellidi per il corpo segmentato, in questo caso rivestito da una cuticola sottile. La testa è ben riconoscibile per la presenza di due antenne. Sul ventre sono presenti numerose appendici locomotorie munite di unghie (in greco, ónicos). I sessi sono separati e il dimorfismo sessuale è accentuato.
Echinodermi
Negli echinodermi (phylum Echinodermata) la larva è a simmetria bilaterale, mentre l’adulto ha simmetria pentaraggiata.
Il corpo è sostenuto da un dermascheletro, cioè un rivestimento di piastre calcaree posto sotto l’epidermide. Non si distingue una vera e propria testa, ma un polo orale, dove si apre la bocca, e un polo aborale, dove si apre l’ano. Tipico degli echinodermi è l’apparato acquifero: questo è formato da un canale circolare posto attorno alla bocca da cui si diramano 5 canali pieni di liquido. In corrispondenza dei canali radiali fuoriescono delle appendici munite di ventose (pedicelli ambulacrali) utilizzate per la locomozione e la respirazione. Quest’ultima funzione è svolta anche, a seconda delle specie, da polmoni o da branchie.
L’apparato circolatorio è formato da lacune parallele ai canali dell’apparato acquifero; un anello posto attorno alla bocca e 5 tronchi radiali formano il sistema nervoso. Anche gli organi riproduttori (gonadi) sono di regola 5.
I sessi sono separati e la riproduzione è sessuata. Lo sviluppo è indiretto con formazione di larve.
| CLASSE | CARATTERISTICHE |
| crinoidi | fissi al substrato marino anche allo stadio larvale, hanno l'aspetto di stelle vivacemente colorate e dotate di uno stelo lungo e sottile (da cui il nome di "gigli di mare"); in alcune specie lo stelo si spezza una volta raggiunta la maturità e gli individui conducono vita libera |
| asteroidi | sono le stelle di mare, con tipica forma a 5 raggi; animali voraci, possono arrivare a distruggere le coltivazioni di molluschi |
| ofiuroidi | sono chiamati stelle serpentine per le braccia sottili e cilindriche e per i caratteristici movimenti con cui strisciano sul fondo |
| echinoidi | sono i ricci di mare, con un corpo globoso rivestito di un esoscheletro rigido provvisto di aculei mobili che servono per la locomozione e la difesa |
| oluturoidi | sono i cetrioli di mare, considerati gli echinodermi più primitivi per la forma a sacco, che maschera la simmetria raggiata |

Un echinoideo o riccio di mare
Cordati
In tutti i cordati (phylum Chordata), nonostante la notevole varietà morfologica, sono presenti le seguenti caratteristiche:
- una corda dorsale (notocorda) di tessuto cartilagineo od osseo, flessibile ed elastica, che fornisce il punto d’attacco per i muscoli
- un tubo neurale, che nella regione cefalica si dilata a formare il cervello e dal quale si dipartono i nervi
- delle fessure branchiali faringee, che possono restare aperte oppure possono chiudersi durante i primi stadi di sviluppo
- una coda che prolunga l’estremità posteriore dello scheletro dell’animale.

Ascidia semplice (Urocordati) |
Amphioxus (Cefalocordati) |

Caracara dalla testa gialla, Milvago chimachima poggiato su un capibara, Hydrochoeris hydrochaeris, gruppo craniata (Emicefalocordati) |
I cordati rappresentano, con gli artropodi e i molluschi, una delle linee evolutive di maggior successo: la ragione di ciò si trova soprattutto nella presenza della notocorda sulla quale si inseriscono i muscoli: questo apparato permette lo sviluppo di movimenti complessi e veloci, come il nuoto, la corsa e il volo.
| CLASSE | RIVESTIMENTO ESTERNO | TEMPERATURA CORPOREA | ARTI | RESPIRAZIONE |
| agnati
o ciclostomi |
pelle viscida | eterotermi | assenza di arti pari | branchie |
| pesci cartilaginei,
o condritti |
scaglie | eterotermi | 2 paia di pinne | branchie |
| pesci ossei,
o osteitti |
scaglie e muco | eterotermi | 2 paia di pinne | branchie |
| anfibi | pelle umida spesso ricoperta di muco | eterotermi | 2 paia di arti; assenza di unghie | branchie; polmoni |
| rettili | pelle secca; squame | eterotermi | 2 paia di arti; unghie | polmoni |
| uccelli | penne; squame sulle zampe | omeotermi | 1 paio di ali; 1 paio di zampe; unghie | polmoni |
| mammiferi,
o mammalia |
peli | omeotermi | 2 paia di zampe; spesso unghie | polmoni |
La sistematica dei cordati si basa sulla posizione della corda: sono considerati più primitivi gli urocordati, in cui la corda è presente solo nella regione caudale della larva, i cefalocordati, che sono un piccolo subphylum posto a cavallo fra gli urocordati e i vertebrati detti anche acranii in quanto privi di cranio a protezione della vescicola encefalica o leptocardii per via della conformazione del cuore, e più evoluti i vertebrati, in cui la corda è circondata o sostituita da uno scheletro cartilagineo od osseo.

Lampreda di fiume, Lampetra fluviatilis (agnata) |

Squalo bianco, Carcharodon carcharias (condritto) |

Tonno, Thunnus (osteitti) |

Rana ibrida dei fossi o rana comune, Pelophylax esculentus (anfibi) |

Tiliqua nigrolutea (rettili) |

Civetta delle nevi, Bubo scandiacus (uccelli) |

Ornitorinco o platipo, Ornithorhynchus anatinus. E' monotremo (solo 5 specie) e depone le uova (mammiferi) |

Topo, Mus musculis (mammiferi) |